

The systematics of the Cervidae: a total evidence approach
- PMID: 32110477
- PMCID: PMC7034380
- DOI: 10.7717/peerj.8114
The systematics of the Cervidae: a total evidence approach
Abstract
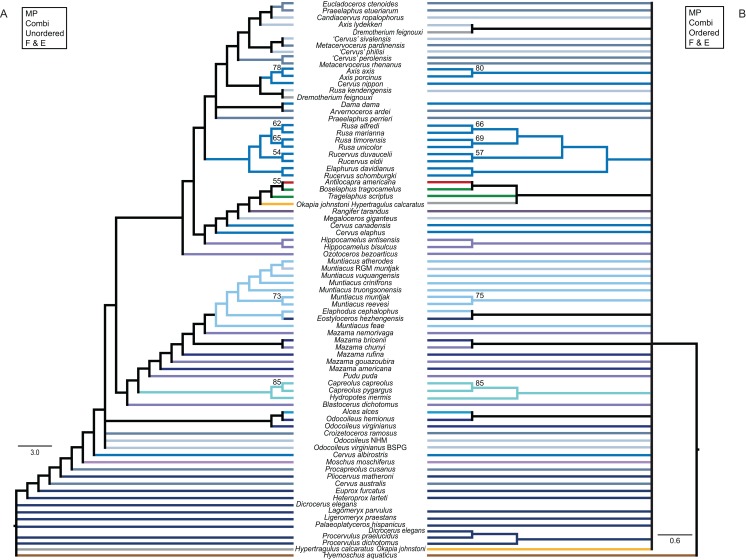
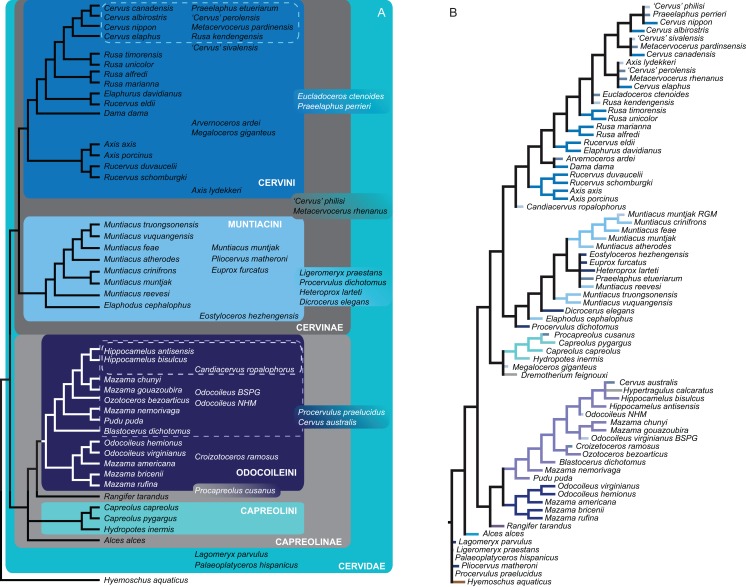
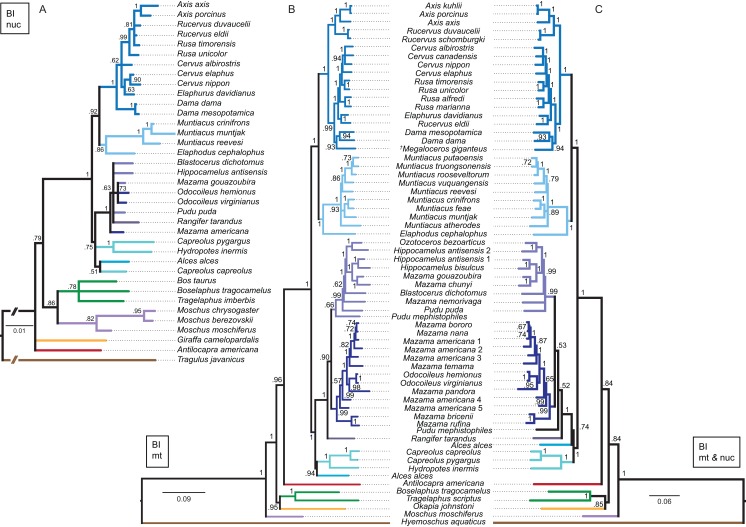
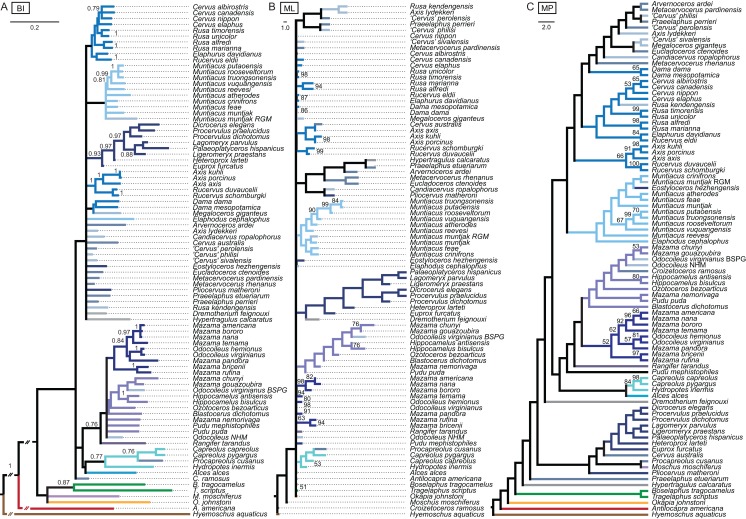
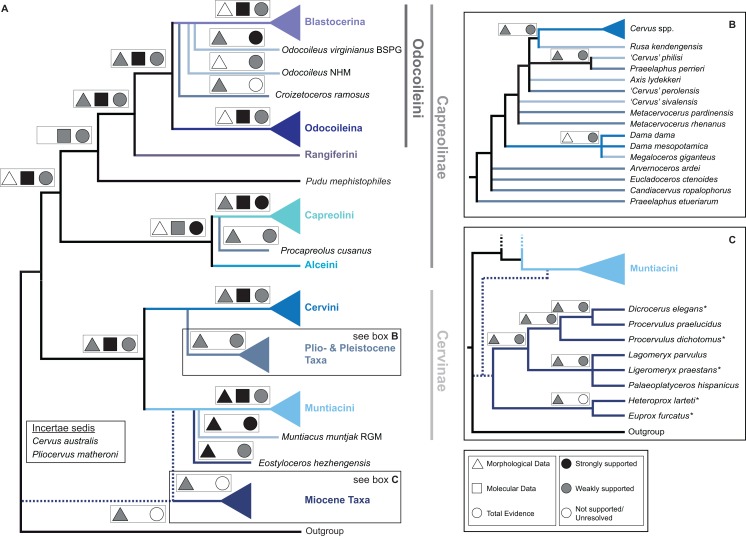
Systematic relationships of cervids have been controversial for decades. Despite new input from molecular systematics, consensus could only be partially reached. The initial, gross (sub) classification based on morphology and comparative anatomy was mostly supported by molecular data. The rich fossil record of cervids has never been extensively tested in phylogenetic frameworks concerning potential systematic relationships of fossil cervids to extant cervids. The aim of this work was to investigate the systematic relationships of extant and fossil cervids using molecular and morphological characters and make implications about their evolutionary history based on the phylogenetic reconstructions. To achieve these objectives, molecular data were compiled consisting of five nuclear markers and the complete mitochondrial genome of 50 extant and one fossil cervids. Several analyses using different data partitions, taxon sampling, partitioning schemes, and optimality criteria were undertaken. In addition, the most extensive morphological character matrix for such a broad cervid taxon sampling was compiled including 168 cranial and dental characters of 41 extant and 29 fossil cervids. The morphological and molecular data were analysed in a combined approach and other comprehensive phylogenetic reconstructions. The results showed that most Miocene cervids were more closely related to each other than to any other cervids. They were often positioned between the outgroup and all other cervids or as the sister taxon to Muntiacini. Two Miocene cervids were frequently placed within Muntiacini. Plio- and Pleistocene cervids could often be affiliated to Cervini, Odocoileini or Capreolini. The phylogenetic analyses provide new insights into the evolutionary history of cervids. Several fossil cervids could be successfully related to living representatives, confirming previously assumed affiliations based on comparative morphology and introducing new hypotheses. New systematic relationships were observed, some uncertainties persisted and resolving systematics within certain taxa remained challenging.
Keywords: Combined analyses; Evolutionary history; Morphology; Phylogeny; Ruminantia.
© 2020 Heckeberg.
Conflict of interest statement
The author declares that she has no competing interests.
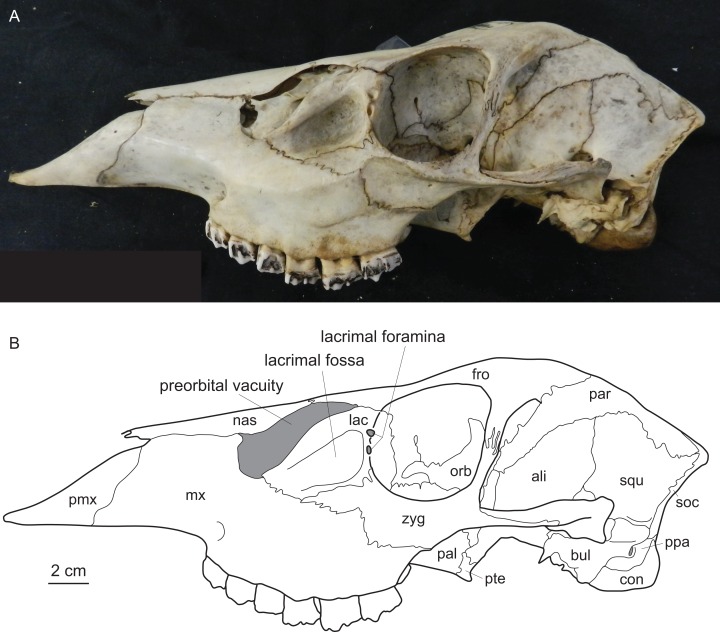
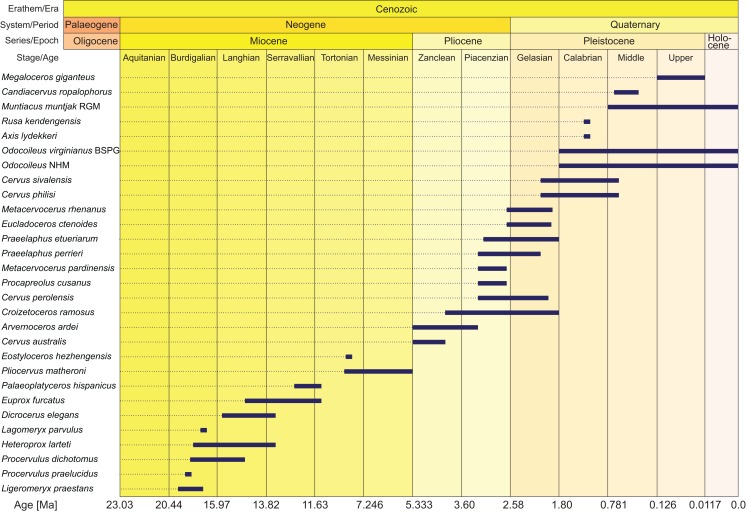
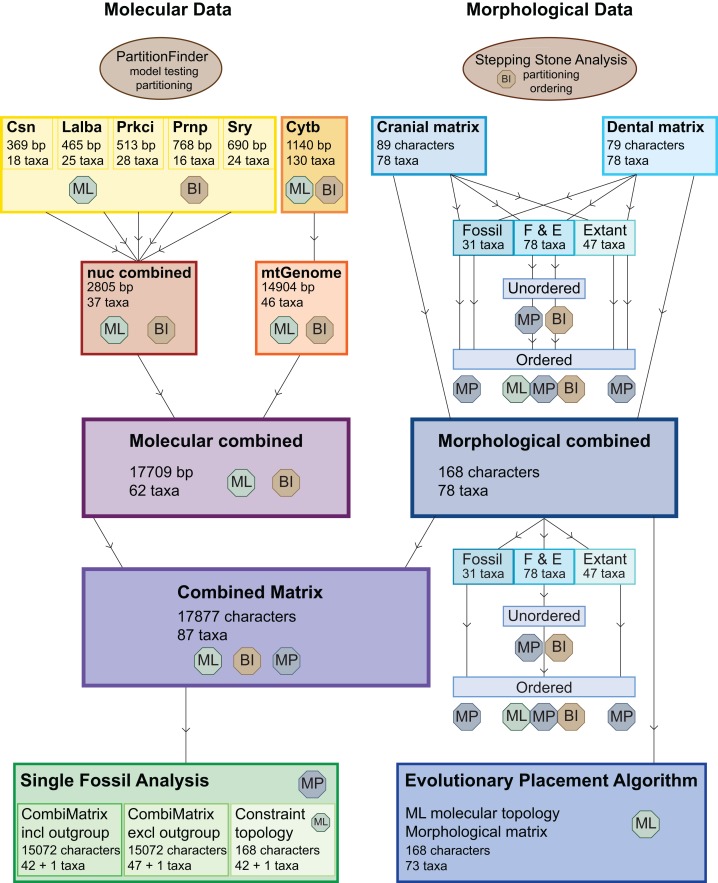
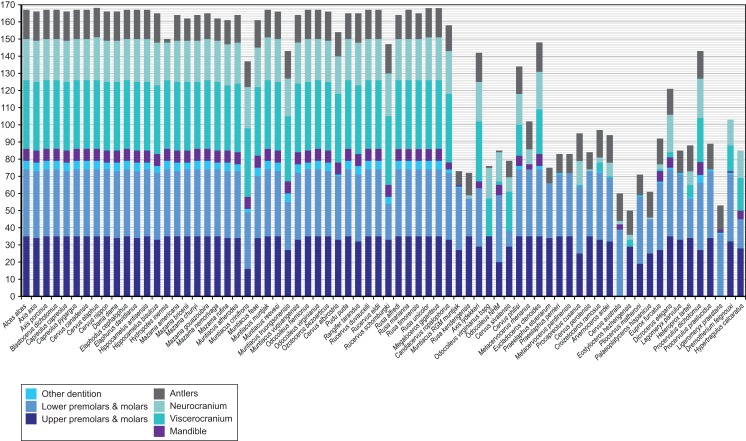
Figures

References
-
- Agnarsson I, May-Collado LJ. The phylogeny of Cetartiodactyla: the importance of dense taxon sampling, missing data, and the remarkable promise of cytochrome b to provide reliable species-level phylogenies. Molecular Phylogenetics and Evolution. 2008;48(3):964–985. doi: 10.1016/j.ympev.2008.05.046. - DOI - PubMed
-
- Alcaraz MA, Zurita AE. Nuevos registros de cérvidos poco conocidos: Epieuryceros cf. proximus Castellanos y Antifer sp. (Mammalia, Artiodactyla, Cervidae) Revista del Museo Argentino de Ciencias Naturales. 2004;6(1):41–48. doi: 10.22179/REVMACN.6.71. - DOI
-
- Amato G, Egan MG, Rabinowitz A. A new species of muntjac, Muntiacus putaoensis (Artiodactyla: Cervidae) from northern Myanmar. Animal Conservation. 1999a;2(1):1–7. doi: 10.1111/j.1469-1795.1999.tb00042.x. - DOI
LinkOut - more resources
Full Text Sources
