

Glycosaminoglycans as Tools to Decipher the Platelet Tumor Cell Interaction: A Focus on P-Selectin
- PMID: 32110917
- PMCID: PMC7179249
- DOI: 10.3390/molecules25051039
Glycosaminoglycans as Tools to Decipher the Platelet Tumor Cell Interaction: A Focus on P-Selectin
Abstract
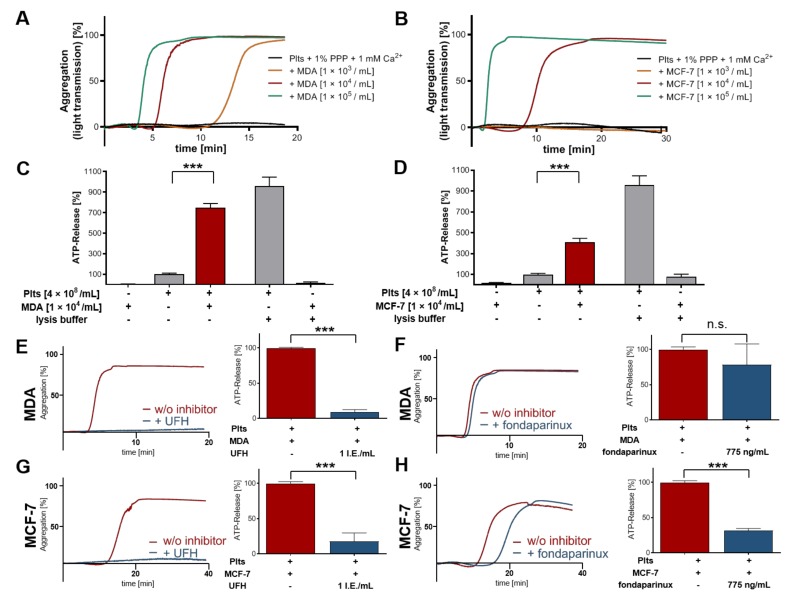
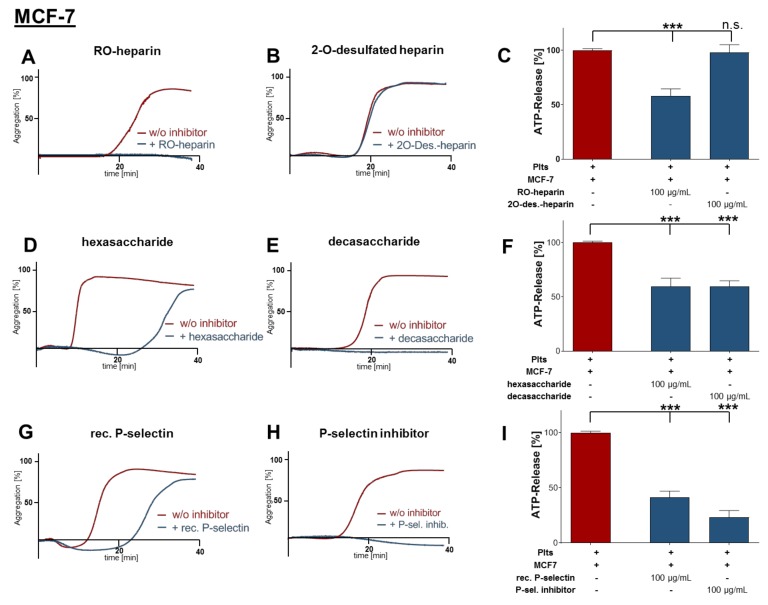
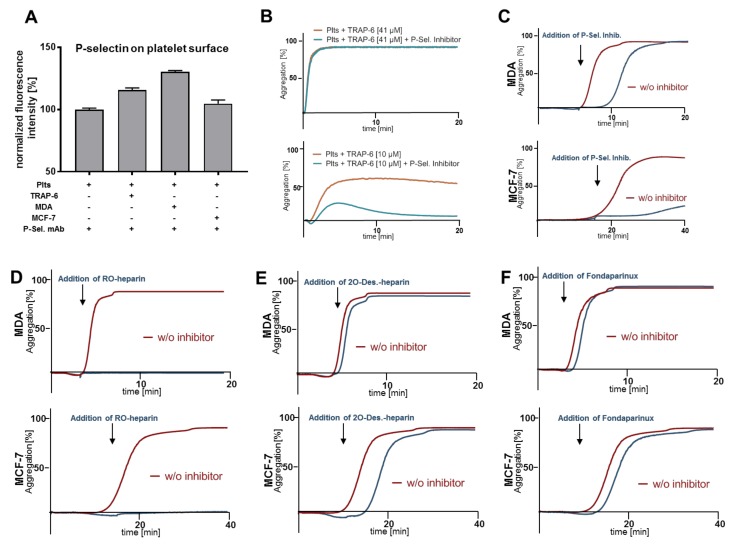
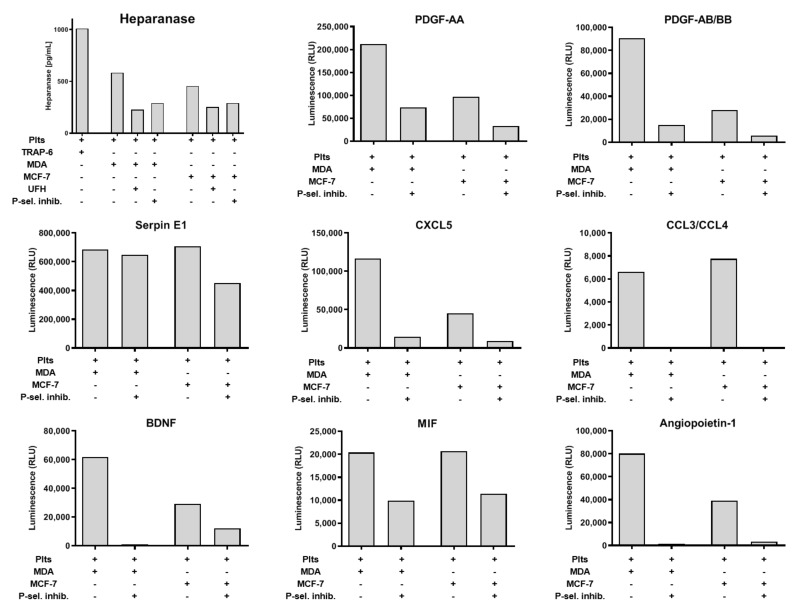
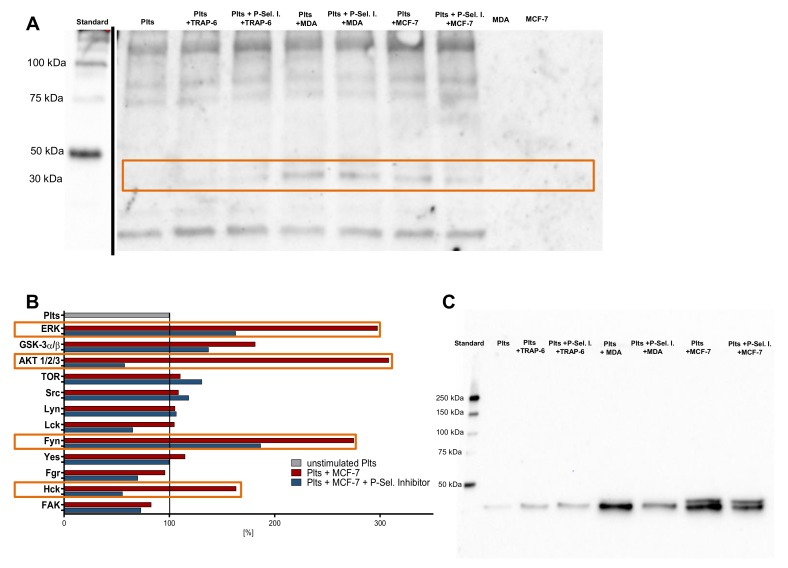
Tumor cell-platelet interactions are regarded as an initial crucial step in hematogenous metastasis. Platelets protect tumor cells from immune surveillance in the blood, mediate vascular arrest, facilitate tumor extravasation, growth, and finally angiogenesis in the metastatic foci. Tumor cells aggregate platelets in the bloodstream by activation of the plasmatic coagulation cascade and by direct contact formation. Antimetastatic activities of unfractionated or low molecular weight heparin (UFH/LMWH) can undoubtedly be related to attenuated platelet activation, but molecular mechanisms and contribution of contact formation vs. coagulation remain to be elucidated. Using a set of non-anticoagulant heparin derivatives varying in size or degree of sulfation as compared with UFH, we provide insight into the relevance of contact formation for platelet activation. Light transmission aggregometry and ATP release assays confirmed that only those heparin derivatives with P-selectin blocking capacities were able to attenuate breast cancer cell-induced platelet activation, while pentasaccharide fondaparinux was without effects. Furthermore, a role of P-selectin in platelet activation and signaling could be confirmed by proteome profiler arrays detecting platelet kinases. In this study, we demonstrate that heparin blocks tumor cell-induced coagulation. Moreover, we identify platelet P-selectin, which obviously acts as molecular switch and controls aggregation and secretion of procoagulant platelets.
Keywords: 2-O-desulfated heparin; P-selectin; RO-heparin; decasaccharide heparin fragment; hexasaccharide heparin fragment; low molecular weight heparin; platelet aggregation; platelet secretion; platelets; tumor metastasis; unfractionated heparin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
The Low Molecular Weight Heparin Tinzaparin Attenuates Platelet Activation in Terms of Metastatic Niche Formation by Coagulation-Dependent and Independent Pathways.Molecules. 2018 Oct 24;23(11):2753. doi: 10.3390/molecules23112753. Molecules. 2018. PMID: 30356007 Free PMC article.
-
A low-anticoagulant heparin suppresses metastatic dissemination through the inhibition of tumor cell-platelets association.Biomed Pharmacother. 2024 Feb;171:116108. doi: 10.1016/j.biopha.2023.116108. Epub 2024 Jan 13. Biomed Pharmacother. 2024. PMID: 38218079
-
Parnaparin, a low-molecular-weight heparin, prevents P-selectin-dependent formation of platelet-leukocyte aggregates in human whole blood.Thromb Haemost. 2007 Jun;97(6):965-73. doi: 10.1160/th06-12-0680. Thromb Haemost. 2007. PMID: 17549299
-
P-selectin, carcinoma metastasis and heparin: novel mechanistic connections with therapeutic implications.Braz J Med Biol Res. 2001 Jun;34(6):711-7. doi: 10.1590/s0100-879x2001000600003. Braz J Med Biol Res. 2001. PMID: 11378658 Review.
-
Heparin inhibition of selectin-mediated interactions during the hematogenous phase of carcinoma metastasis: rationale for clinical studies in humans.Semin Thromb Hemost. 2002 Feb;28(1):53-66. doi: 10.1055/s-2002-20564. Semin Thromb Hemost. 2002. PMID: 11885026 Review.
Cited by
-
Doxorubicin-Loaded Platelet Decoys for Enhanced Chemoimmunotherapy Against Triple-Negative Breast Cancer in Mice Model.Int J Nanomedicine. 2023 Jun 30;18:3577-3593. doi: 10.2147/IJN.S403339. eCollection 2023. Int J Nanomedicine. 2023. PMID: 37409026 Free PMC article.
-
Shear Stress Accumulation Enhances von Willebrand Factor-Induced Platelet P-Selectin Translocation in a PI3K/Akt Pathway-Dependent Manner.Front Cell Dev Biol. 2021 Jun 1;9:642108. doi: 10.3389/fcell.2021.642108. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34141704 Free PMC article.
-
Thrombus-Targeting Polymeric Nanocarriers and Their Biomedical Applications in Thrombolytic Therapy.Front Physiol. 2021 Nov 30;12:763085. doi: 10.3389/fphys.2021.763085. eCollection 2021. Front Physiol. 2021. PMID: 34916956 Free PMC article. Review.
-
Exostosin 1 Knockdown Induces Chemoresistance in MV3 Melanoma Cells by Upregulating JNK and MEK/ERK Signaling.Int J Mol Sci. 2023 Mar 13;24(6):5452. doi: 10.3390/ijms24065452. Int J Mol Sci. 2023. PMID: 36982528 Free PMC article.
-
A Combined Activity of Thrombin and P-Selectin Is Essential for Platelet Activation by Pancreatic Cancer Cells.Int J Mol Sci. 2021 Mar 24;22(7):3323. doi: 10.3390/ijms22073323. Int J Mol Sci. 2021. PMID: 33805059 Free PMC article.
References
-
- Labelle M., Hynes R.O. The initial hours of metastasis: The importance of cooperative host-tumor cell interactions during hematogenous dissemination. Cancer Discov. 2012;2:1091–1099. doi: 10.1158/2159-8290.CD-12-0329. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
