

The microtubule-associated histone methyltransferase SET8, facilitated by transcription factor LSF, methylates α-tubulin
- PMID: 32111740
- PMCID: PMC7135998
- DOI: 10.1074/jbc.RA119.010951
The microtubule-associated histone methyltransferase SET8, facilitated by transcription factor LSF, methylates α-tubulin
Abstract
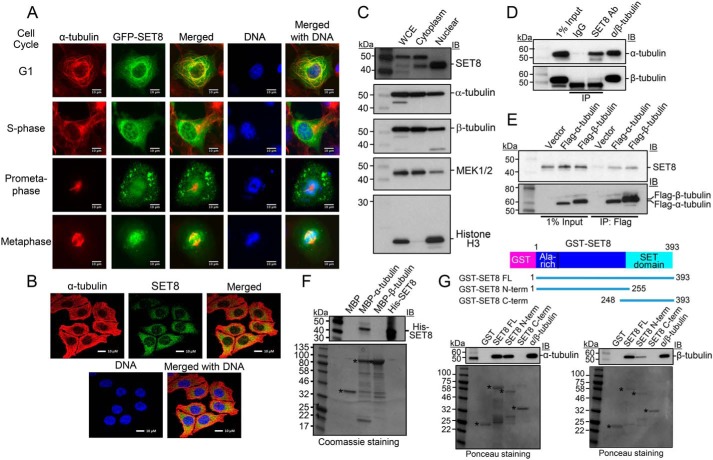
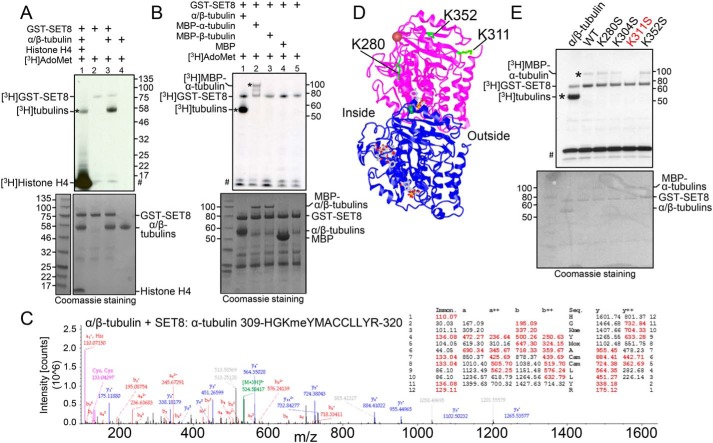
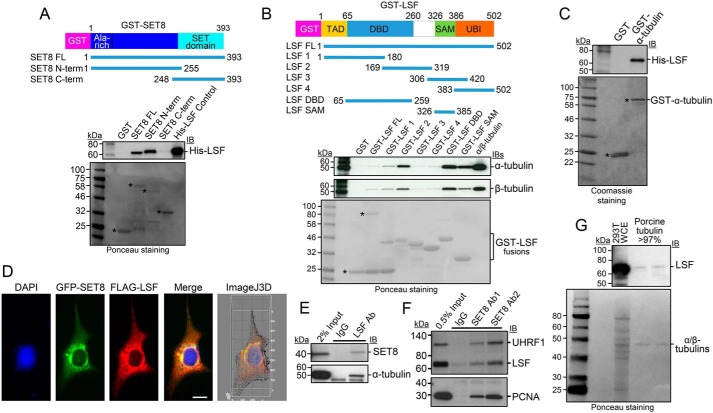
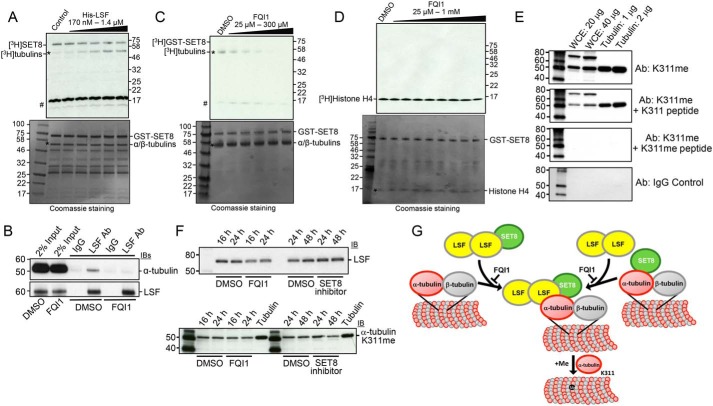
Microtubules are cytoskeletal structures critical for mitosis, cell motility, and protein and organelle transport and are a validated target for anticancer drugs. However, how tubulins are regulated and recruited to support these distinct cellular processes is incompletely understood. Posttranslational modifications of tubulins are proposed to regulate microtubule function and dynamics. Although many of these modifications have been investigated, only one prior study reports tubulin methylation and an enzyme responsible for this methylation. Here we used in vitro radiolabeling, MS, and immunoblotting approaches to monitor protein methylation and immunoprecipitation, immunofluorescence, and pulldown approaches to measure protein-protein interactions. We demonstrate that N-lysine methyltransferase 5A (KMT5A or SET8/PR-Set7), which methylates lysine 20 in histone H4, bound α-tubulin and methylated it at a specific lysine residue, Lys311 Furthermore, late SV40 factor (LSF)/CP2, a known transcription factor, bound both α-tubulin and SET8 and enhanced SET8-mediated α-tubulin methylation in vitro In addition, we found that the ability of LSF to facilitate this methylation is countered by factor quinolinone inhibitor 1 (FQI1), a specific small-molecule inhibitor of LSF. These findings suggest the general model that microtubule-associated proteins, including transcription factors, recruit or stimulate protein-modifying enzymes to target tubulins. Moreover, our results point to dual functions for SET8 and LSF not only in chromatin regulation but also in cytoskeletal modification.
Keywords: SET8; cancer; cytoskeleton; lysine methyltransferase 5A (KMT5A); mammalian cells; posttranslational modification (PTM); protein methylation; transcription factor CP2 (TFCP2); transcription factor LSF; tubulin.
© 2020 Chin et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
