

Pharmacological LRH-1/Nr5a2 inhibition limits pro-inflammatory cytokine production in macrophages and associated experimental hepatitis
- PMID: 32111818
- PMCID: PMC7048823
- DOI: 10.1038/s41419-020-2348-9
Pharmacological LRH-1/Nr5a2 inhibition limits pro-inflammatory cytokine production in macrophages and associated experimental hepatitis
Abstract
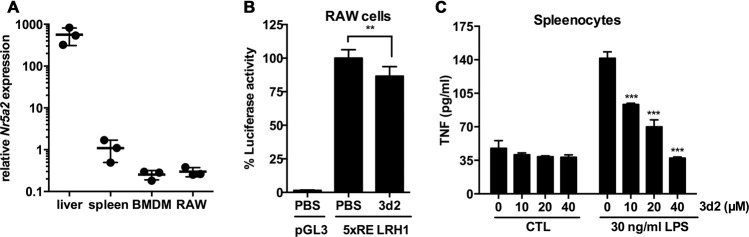
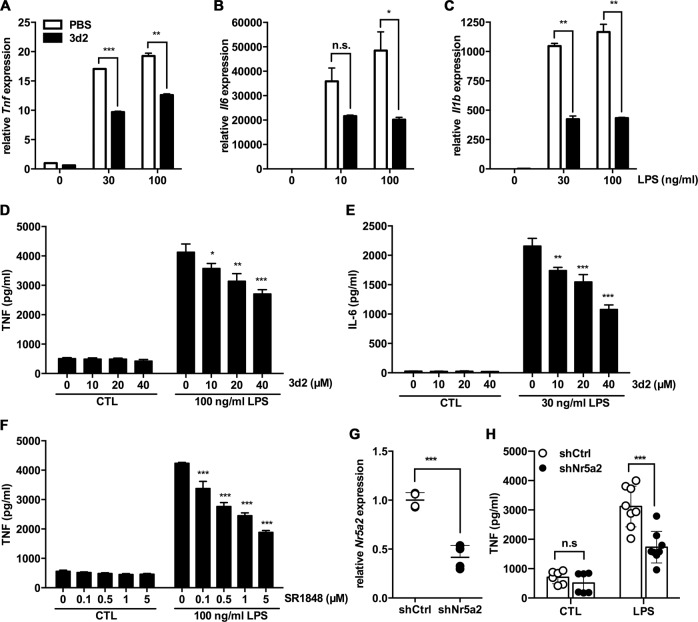
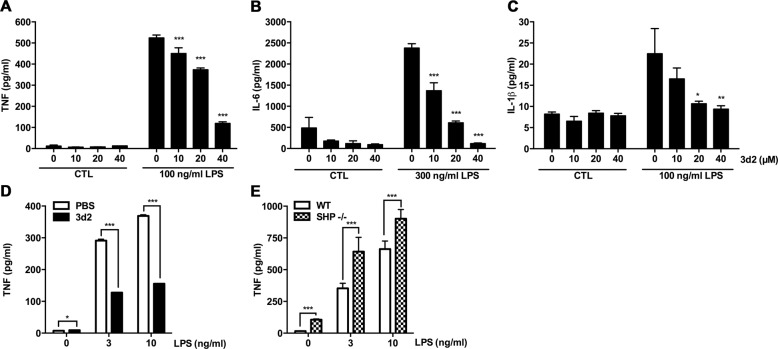
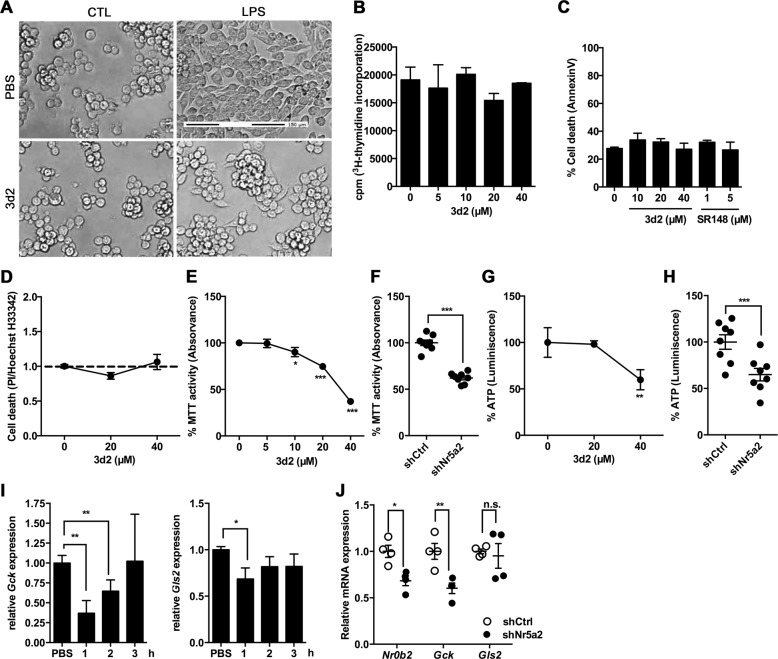
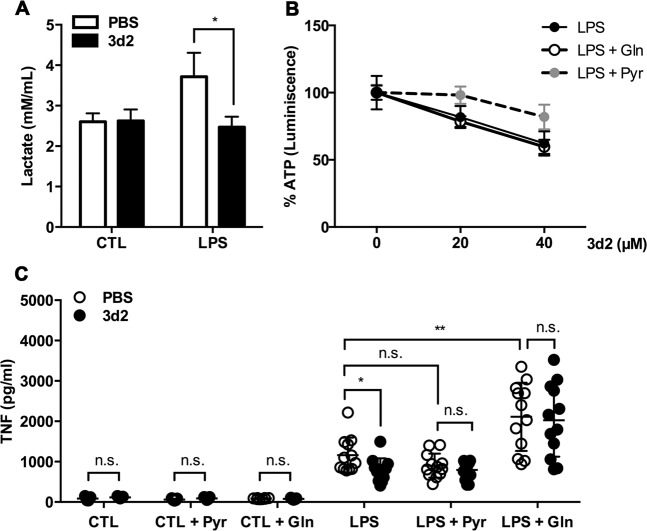
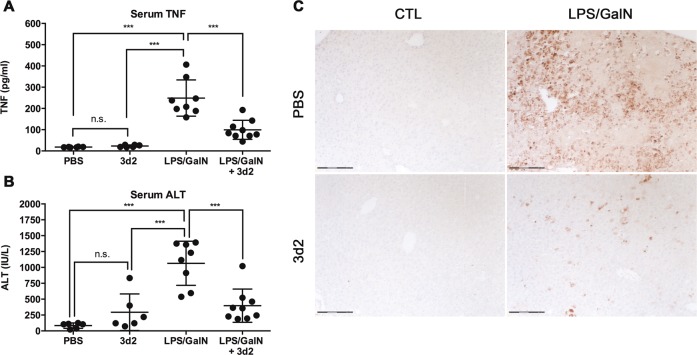
Liver receptor homolog-1 (LRH-1, Nr5a2) is an orphan nuclear receptor mainly expressed in tissues of endodermal origin, where its physiological role has been extensively studied. LRH-1 has been implicated in liver cell differentiation and proliferation, as well as glucose, lipid, and bile acid metabolism. In addition, increasing evidence highlights its role in immunoregulatory processes via glucocorticoid synthesis in the intestinal epithelium. Although the direct function of LRH-1 in immune cells is fairly elucidated, a role of LRH-1 in the regulation of macrophage differentiation has been recently reported. In this study, we aimed to investigate the role of LRH-1 in the regulation of pro-inflammatory cytokine production in macrophages. Our data demonstrate that pharmacological inhibition, along with LRH-1 knockdown, significantly reduced the lipopolysaccharide (LPS)-induced production of pro-inflammatory cytokines in the macrophage line RAW 264.7 cells, as well as in primary murine macrophages. This inhibitory effect was found to be independent of defects of LRH-1-regulated cell proliferation or toxic effects of the LRH-1 inhibitors. In contrast, LRH-1 inhibition reduced the mitochondrial ATP production and metabolism of macrophages through downregulation of the LRH-1 targets glucokinase and glutminase-2, and thus impairing the LPS-induced macrophage activation. Interestingly, in vivo pharmacological inhibition of LRH-1 also resulted in reduced tumor necrosis factor (TNF) production and associated decreased liver damage in a macrophage- and TNF-dependent mouse model of hepatitis. Noteworthy, despite hepatocytes expressing high levels of LRH-1, pharmacological inhibition of LRH-1 per se did not cause any obvious liver damage. Therefore, this study proposes LRH-1 as an emerging therapeutic target in the treatment of inflammatory disorders, especially where macrophages and cytokines critically decide the extent of inflammation.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
