

Pores for Thought: Can Genetic Manipulation of Stomatal Density Protect Future Rice Yields?
- PMID: 32117345
- PMCID: PMC7026486
- DOI: 10.3389/fpls.2019.01783
Pores for Thought: Can Genetic Manipulation of Stomatal Density Protect Future Rice Yields?
Abstract
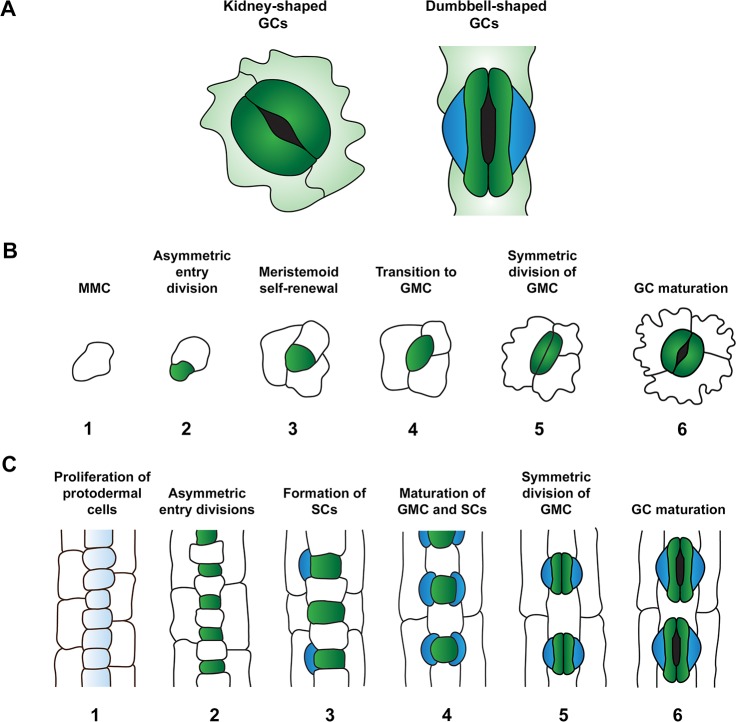
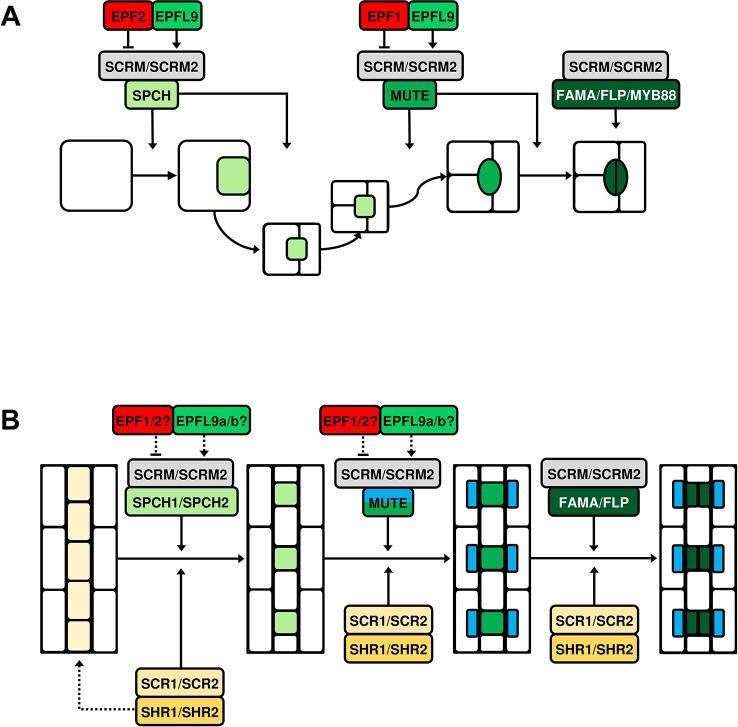
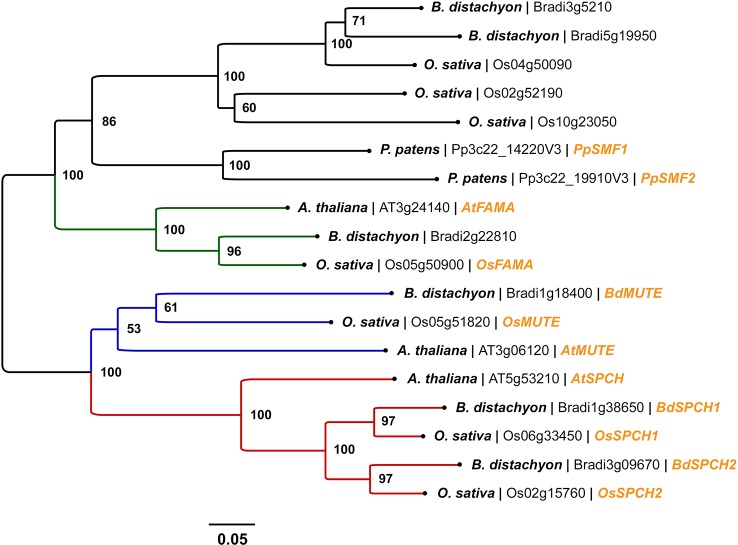
Rice (Oryza sativa L.) contributes to the diets of around 3.5 billion people every day and is consumed more than any other plant. Alarmingly, climate predictions suggest that the frequency of severe drought and high-temperature events will increase, and this is set to threaten the global rice supply. In this review, we consider whether water or heat stresses in crops - especially rice - could be mitigated through alterations to stomata; minute pores on the plant epidermis that permit carbon acquisition and regulate water loss. In the short-term, water loss is controlled via alterations to the degree of stomatal "openness", or, in the longer-term, by altering the number (or density) of stomata that form. A range of molecular components contribute to the regulation of stomatal density, including transcription factors, plasma membrane-associated proteins and intercellular and extracellular signaling molecules. Much of our existing knowledge relating to stomatal development comes from research conducted on the model plant, Arabidopsis thaliana. However, due to the importance of cereal crops to global food supply, studies on grass stomata have expanded in recent years, with molecular-level discoveries underscoring several divergent developmental and morphological features. Cultivation of rice is particularly water-intensive, and there is interest in developing varieties that require less water yet still maintain grain yields. This could be achieved by manipulating stomatal development; a crop with fewer stomata might be more conservative in its water use and therefore more capable of surviving periods of water stress. However, decreasing stomatal density might restrict the rate of CO2 uptake and reduce the extent of evaporative cooling, potentially leading to detrimental effects on yields. Thus, the extent to which crop yields in the future climate will be affected by increasing or decreasing stomatal density should be determined. Here, our current understanding of the regulation of stomatal development is summarised, focusing particularly on the genetic mechanisms that have recently been described for rice and other grasses. Application of this knowledge toward the creation of "climate-ready" rice is discussed, with attention drawn to the lesser-studied molecular elements whose contributions to the complexity of grass stomatal development must be understood to advance efforts.
Keywords: crop physiology; crop yields; grasses; rice; stomatal density; stomatal development.
Copyright © 2020 Buckley, Caine and Gray.
Figures
References
-
- Arve L., Torre S., Olsen J., Tanino K. (2011). “Stomatal Responses to Drought Stress and Air Humidity,” in Abiotic Stress in Plants - Mechanisms and Adaptations (Rijeka: IntechOpen; ), 268–280.
Publication types
LinkOut - more resources
Full Text Sources