

How to Inactivate Human Ubiquitin E3 Ligases by Mutation
- PMID: 32117970
- PMCID: PMC7010608
- DOI: 10.3389/fcell.2020.00039
How to Inactivate Human Ubiquitin E3 Ligases by Mutation
Abstract
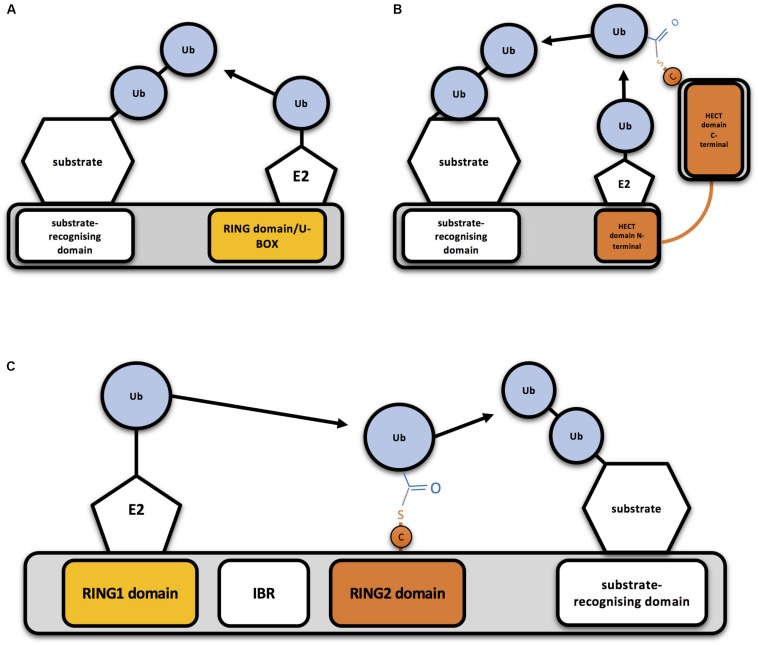
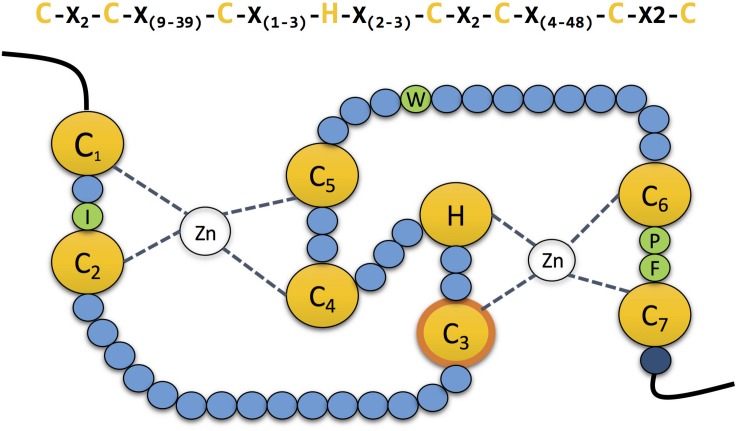
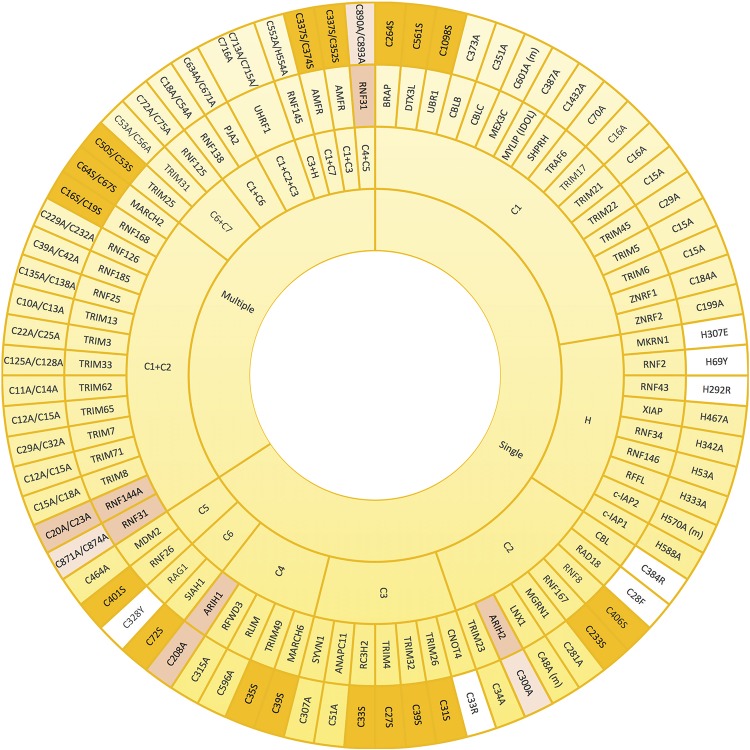
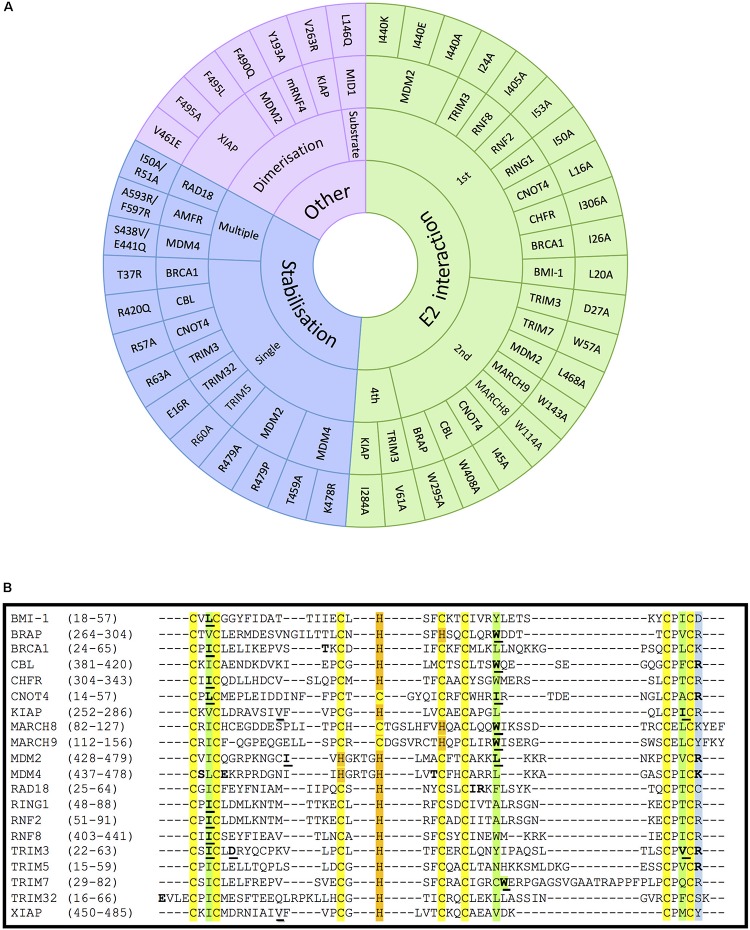
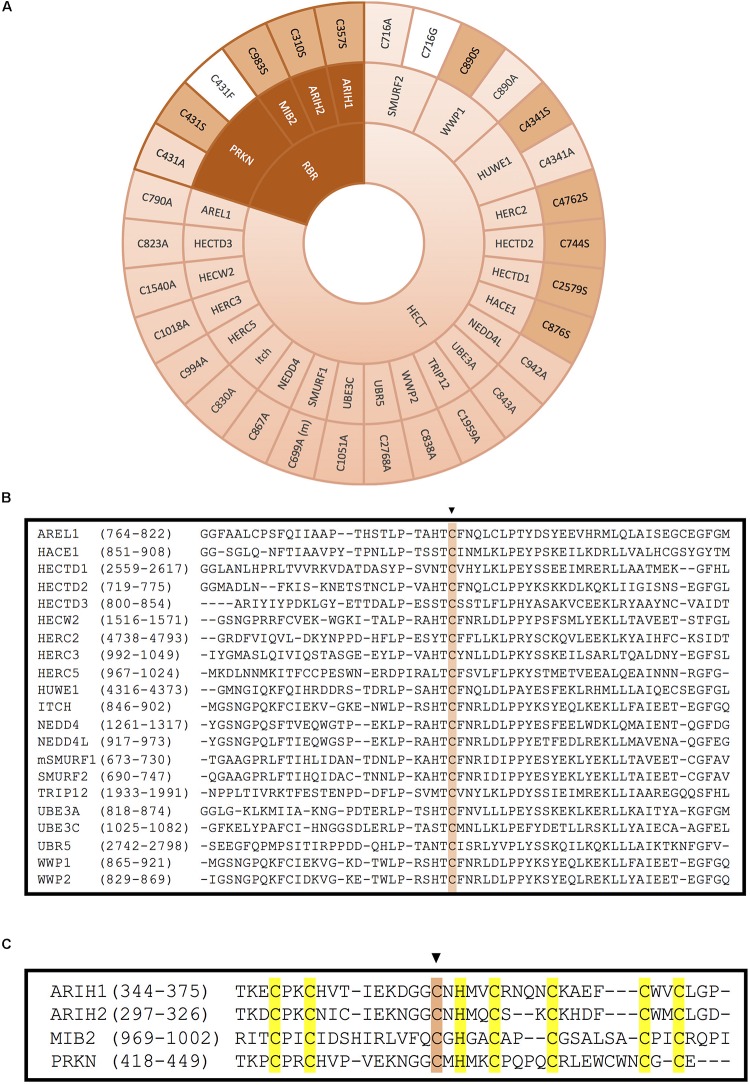
E3 ubiquitin ligases are the ultimate enzymes involved in the transfer of ubiquitin to substrate proteins, a process that determines the fate of the modified protein. Numerous diseases are caused by defects in the ubiquitin-proteasome machinery, including when the activity of a given E3 ligase is hampered. Thus, inactivation of E3 ligases and the resulting effects at molecular or cellular level have been the focus of many studies during the last few years. For this purpose, site-specific mutation of key residues involved in either protein interaction, substrate recognition or ubiquitin transfer have been reported to successfully inactivate E3 ligases. Nevertheless, it is not always trivial to predict which mutation(s) will block the catalytic activity of a ligase. Here we review over 250 site-specific inactivating mutations that have been carried out in 120 human E3 ubiquitin ligases. We foresee that the information gathered here will be helpful for the design of future experimental strategies.
Keywords: E3; inactivation; ligase; mutation; ubiquitin.
Copyright © 2020 Garcia-Barcena, Osinalde, Ramirez and Mayor.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources