

Physical interactions between Gsx2 and Ascl1 balance progenitor expansion versus neurogenesis in the mouse lateral ganglionic eminence
- PMID: 32122989
- PMCID: PMC7157596
- DOI: 10.1242/dev.185348
Physical interactions between Gsx2 and Ascl1 balance progenitor expansion versus neurogenesis in the mouse lateral ganglionic eminence
Abstract
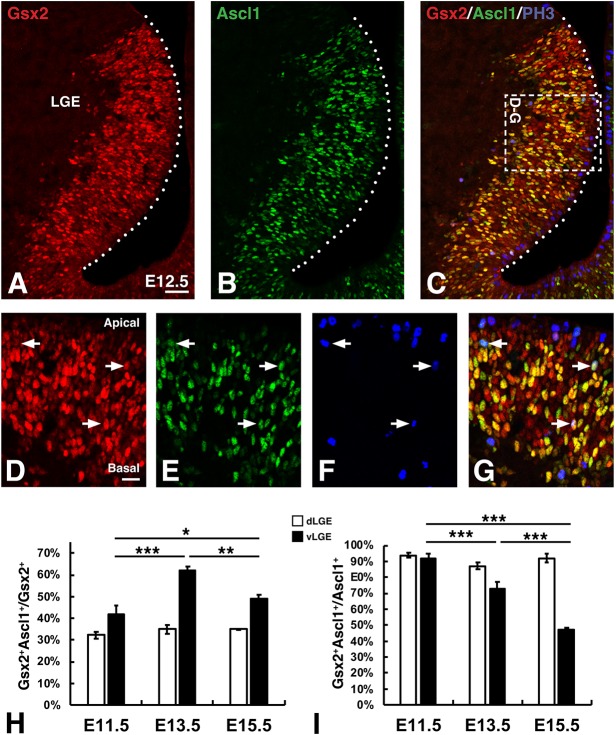
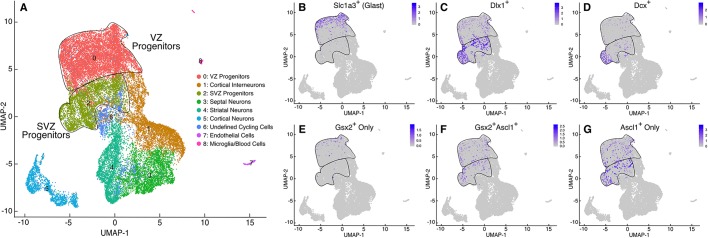
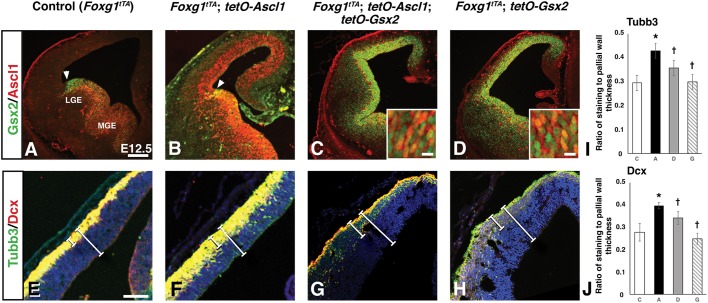
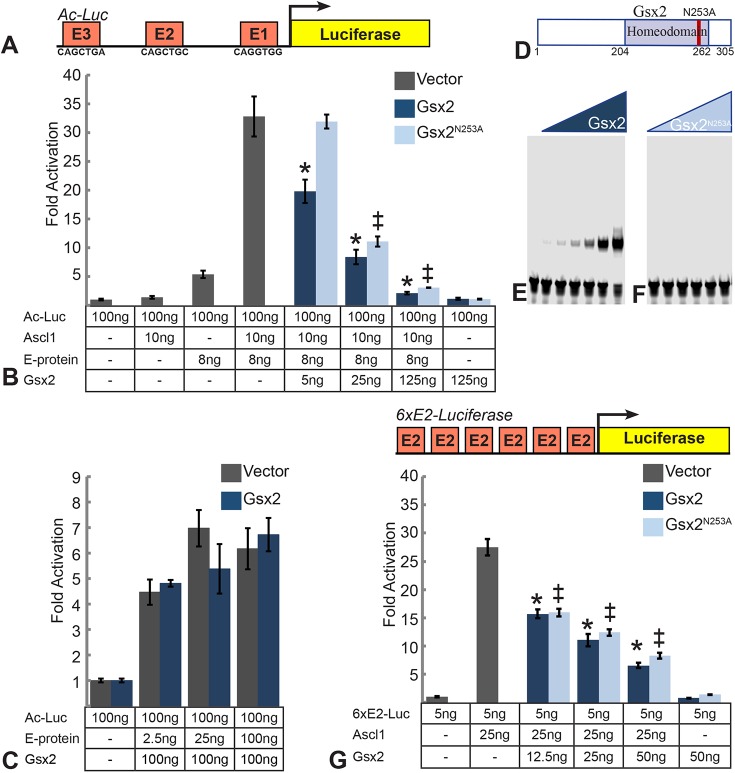
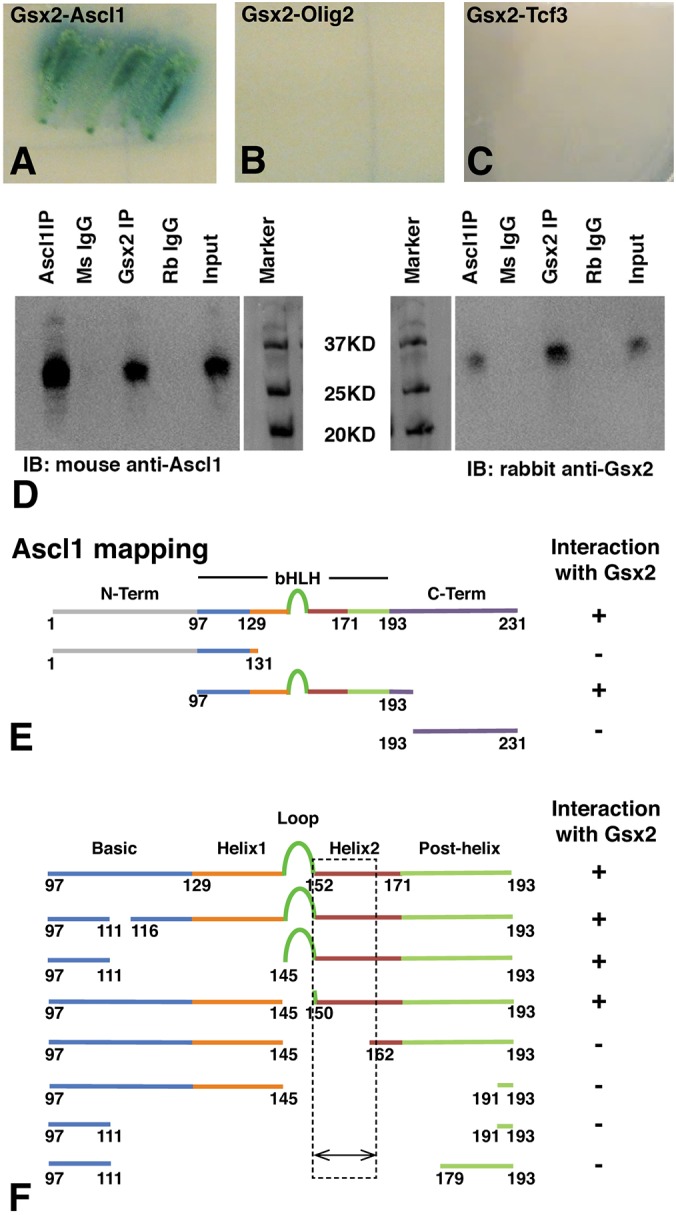
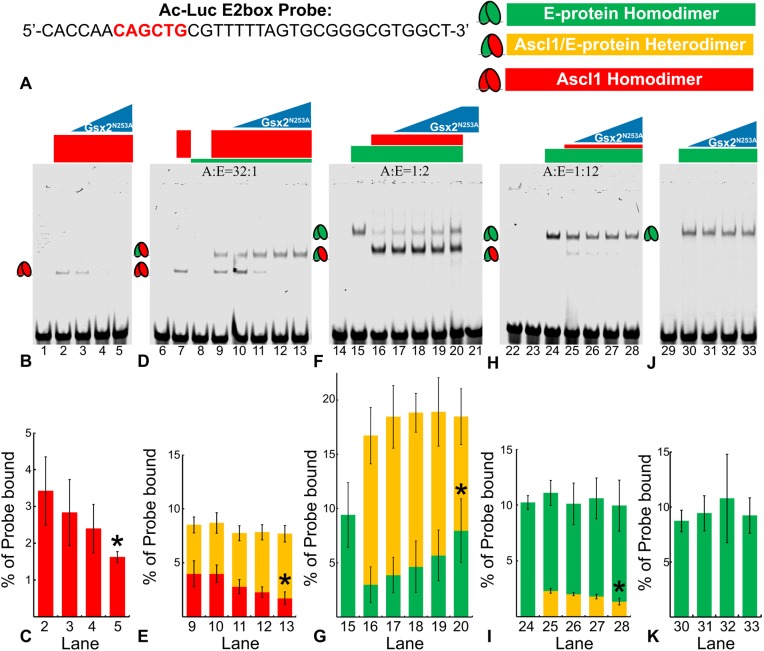
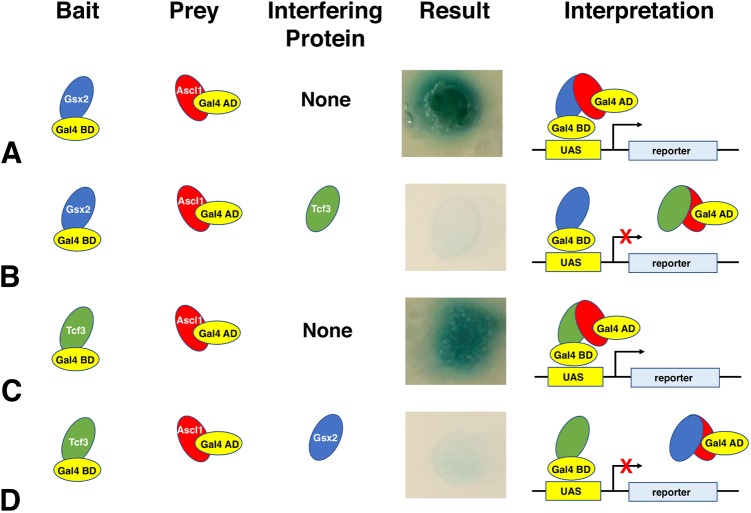
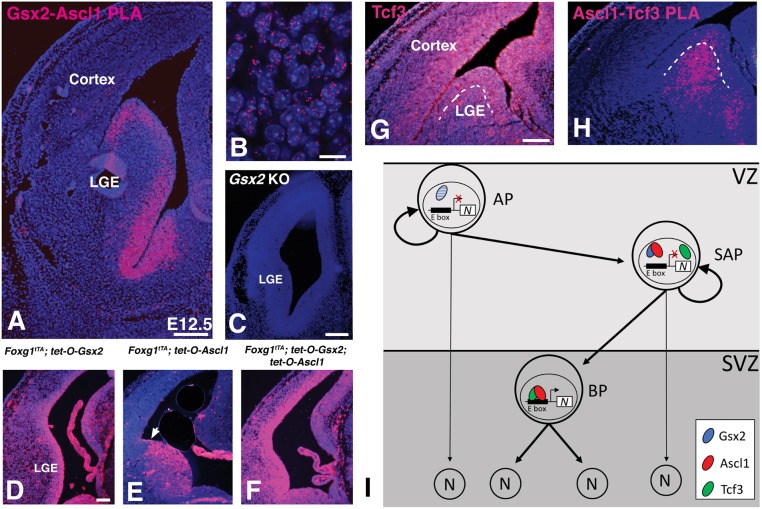
The Gsx2 homeodomain transcription factor promotes neural progenitor identity in the lateral ganglionic eminence (LGE), despite upregulating the neurogenic factor Ascl1. How this balance in maturation is maintained is unclear. Here, we show that Gsx2 and Ascl1 are co-expressed in subapical progenitors that have unique transcriptional signatures in LGE ventricular zone (VZ) cells. Moreover, whereas Ascl1 misexpression promotes neurogenesis in dorsal telencephalic progenitors, the co-expression of Gsx2 with Ascl1 inhibits neurogenesis. Using luciferase assays, we found that Gsx2 reduces the ability of Ascl1 to activate gene expression in a dose-dependent and DNA binding-independent manner. Furthermore, Gsx2 physically interacts with the basic helix-loop-helix (bHLH) domain of Ascl1, and DNA-binding assays demonstrated that this interaction interferes with the ability of Ascl1 to bind DNA. Finally, we modified a proximity ligation assay for tissue sections and found that Ascl1-Gsx2 interactions are enriched within LGE VZ progenitors, whereas Ascl1-Tcf3 (E-protein) interactions predominate in the subventricular zone. Thus, Gsx2 contributes to the balance between progenitor maintenance and neurogenesis by physically interacting with Ascl1, interfering with its DNA binding and limiting neurogenesis within LGE progenitors.
Keywords: E-protein; Proximity ligation assay; Subapical progenitor; Tcf3; Telencephalon.
© 2020. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
Similar articles
-
Gsx transcription factors control neuronal versus glial specification in ventricular zone progenitors of the mouse lateral ganglionic eminence.Dev Biol. 2018 Oct 1;442(1):115-126. doi: 10.1016/j.ydbio.2018.07.005. Epub 2018 Jul 7. Dev Biol. 2018. PMID: 29990475 Free PMC article.
-
Ascl1 is a required downstream effector of Gsx gene function in the embryonic mouse telencephalon.Neural Dev. 2009 Feb 10;4:5. doi: 10.1186/1749-8104-4-5. Neural Dev. 2009. PMID: 19208224 Free PMC article.
-
The homeobox gene Gsx2 controls the timing of oligodendroglial fate specification in mouse lateral ganglionic eminence progenitors.Development. 2013 Jun;140(11):2289-98. doi: 10.1242/dev.091090. Epub 2013 May 1. Development. 2013. PMID: 23637331 Free PMC article.
-
Old and new functions of proneural factors revealed by the genome-wide characterization of their transcriptional targets.Cell Cycle. 2011 Dec 1;10(23):4026-31. doi: 10.4161/cc.10.23.18578. Epub 2011 Dec 1. Cell Cycle. 2011. PMID: 22101262 Free PMC article. Review.
-
Proneural genes in neocortical development.Neuroscience. 2013 Dec 3;253:256-73. doi: 10.1016/j.neuroscience.2013.08.029. Epub 2013 Aug 30. Neuroscience. 2013. PMID: 23999125 Review.
Cited by
-
Single cell enhancer activity distinguishes GABAergic and cholinergic lineages in embryonic mouse basal ganglia.Proc Natl Acad Sci U S A. 2022 Apr 12;119(15):e2108760119. doi: 10.1073/pnas.2108760119. Epub 2022 Apr 4. Proc Natl Acad Sci U S A. 2022. PMID: 35377797 Free PMC article.
-
From Progenitors to Progeny: Shaping Striatal Circuit Development and Function.J Neurosci. 2021 Nov 17;41(46):9483-9502. doi: 10.1523/JNEUROSCI.0620-21.2021. J Neurosci. 2021. PMID: 34789560 Free PMC article. Review.
-
FGF8-FGFR1 signaling regulates human GnRH neuron differentiation in a time- and dose-dependent manner.Dis Model Mech. 2022 Aug 1;15(8):dmm049436. doi: 10.1242/dmm.049436. Epub 2022 Aug 16. Dis Model Mech. 2022. PMID: 35833364 Free PMC article.
-
Conserved Gsx2/Ind homeodomain monomer versus homodimer DNA binding defines regulatory outcomes in flies and mice.Genes Dev. 2021 Jan 1;35(1-2):157-174. doi: 10.1101/gad.343053.120. Epub 2020 Dec 17. Genes Dev. 2021. PMID: 33334823 Free PMC article.
-
Modelling a pathological GSX2 variant that selectively alters DNA binding reveals hypomorphic mouse brain defects.Dis Model Mech. 2025 Feb 1;18(2):dmm052110. doi: 10.1242/dmm.052110. Epub 2025 Feb 20. Dis Model Mech. 2025. PMID: 39882631 Free PMC article.
References
-
- Berger M. F., Badis G., Gehrke A. R., Talukder S., Philippakis A. A., Peña-Castillo L., Alleyne T. M., Mnaimenh S., Botvinnik O. B., Chan E. T. et al. (2008). Variation in homeodomain DNA binding revealed by high-resolution analysis of sequence preferences. Cell 133, 1266-1276. 10.1016/j.cell.2008.05.024 - DOI - PMC - PubMed
-
- Castro D. S., Martynoga B., Parras C., Ramesh V., Pacary E., Johnston C., Drechsel D., Lebel-Potter M., Garcia L. G., Hunt C. et al. (2011). A novel function of the proneural factor Ascl1 in progenitor proliferation identified by genome-wide characterization of its targets. Genes. Dev. 25, 930-945. 10.1101/gad.627811 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
