

Single-cell analysis of human ovarian cortex identifies distinct cell populations but no oogonial stem cells
- PMID: 32123174
- PMCID: PMC7052271
- DOI: 10.1038/s41467-020-14936-3
Single-cell analysis of human ovarian cortex identifies distinct cell populations but no oogonial stem cells
Abstract
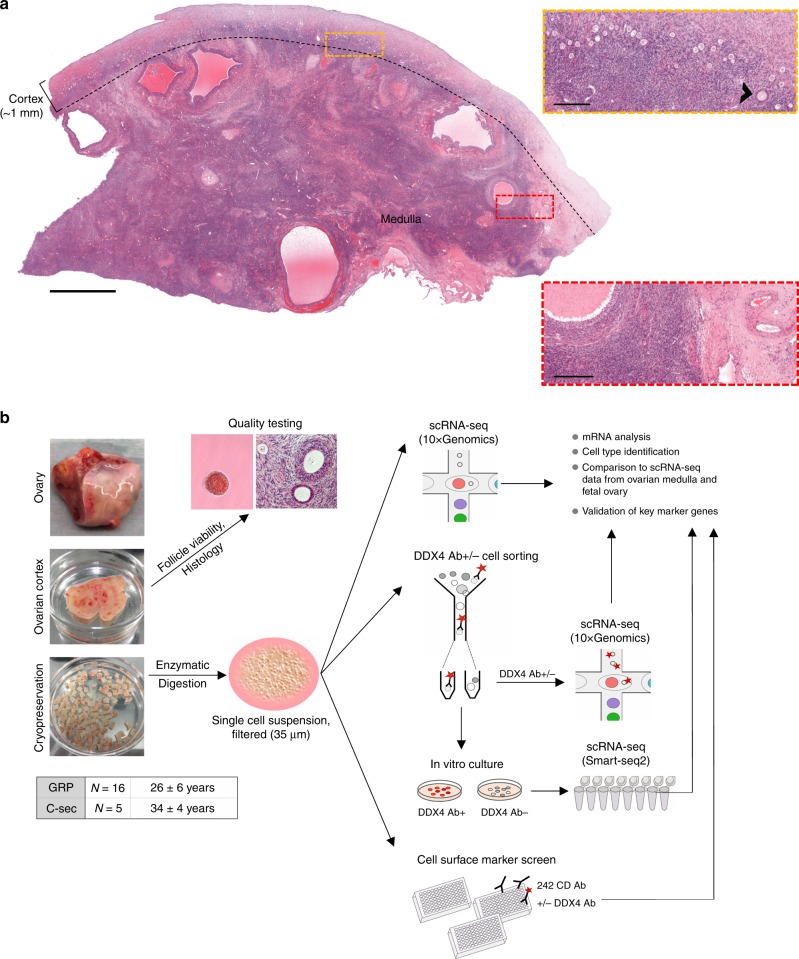
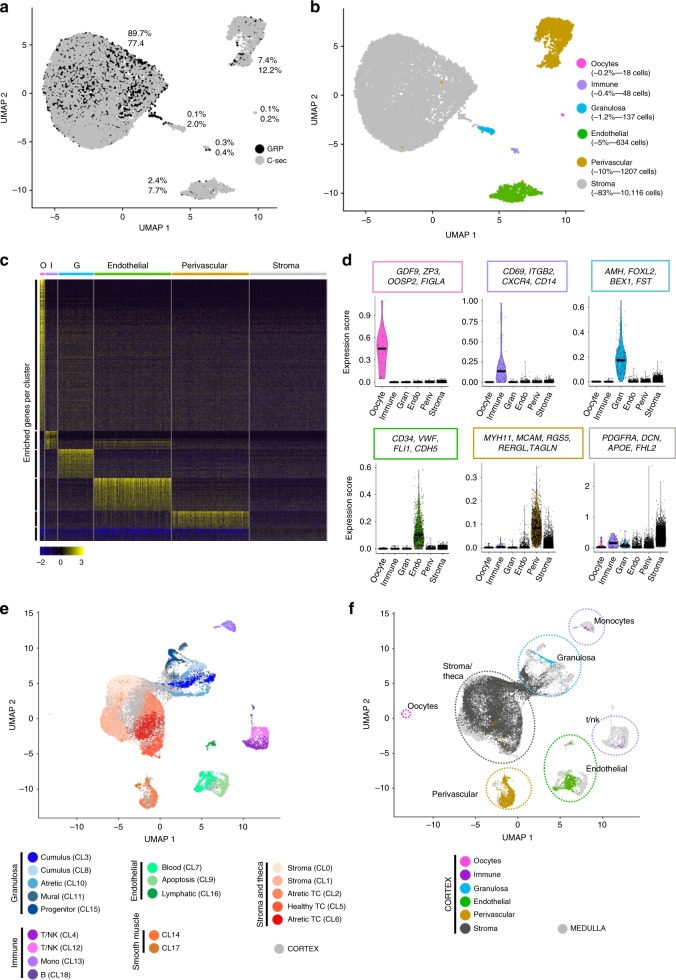
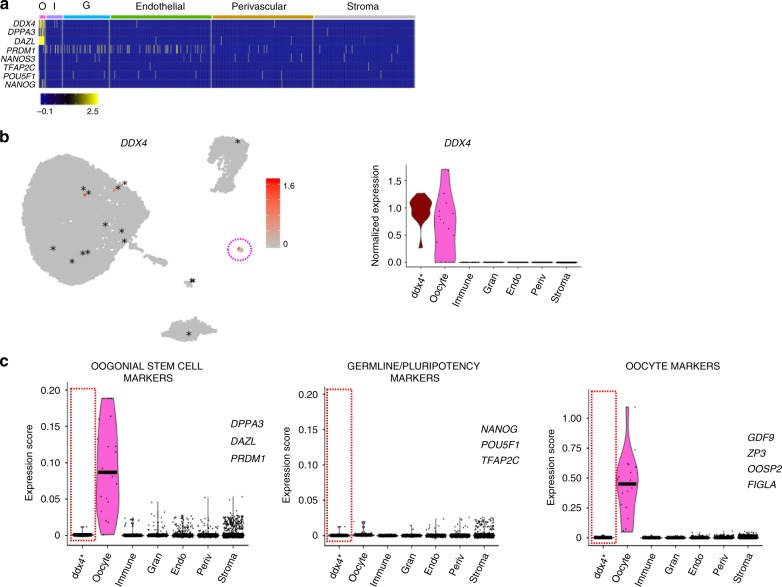
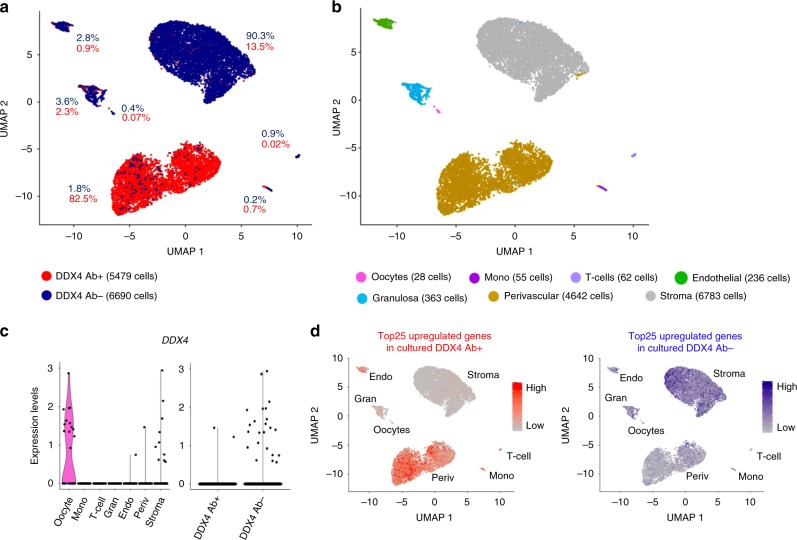
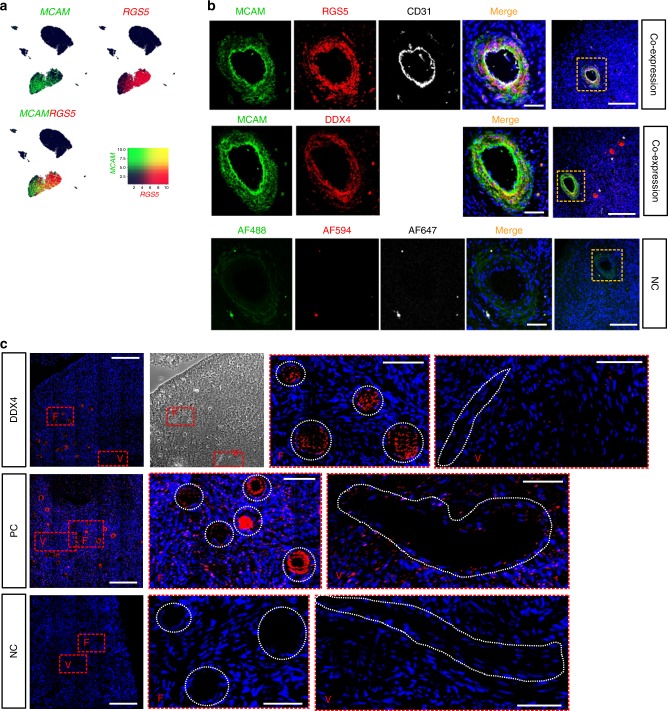
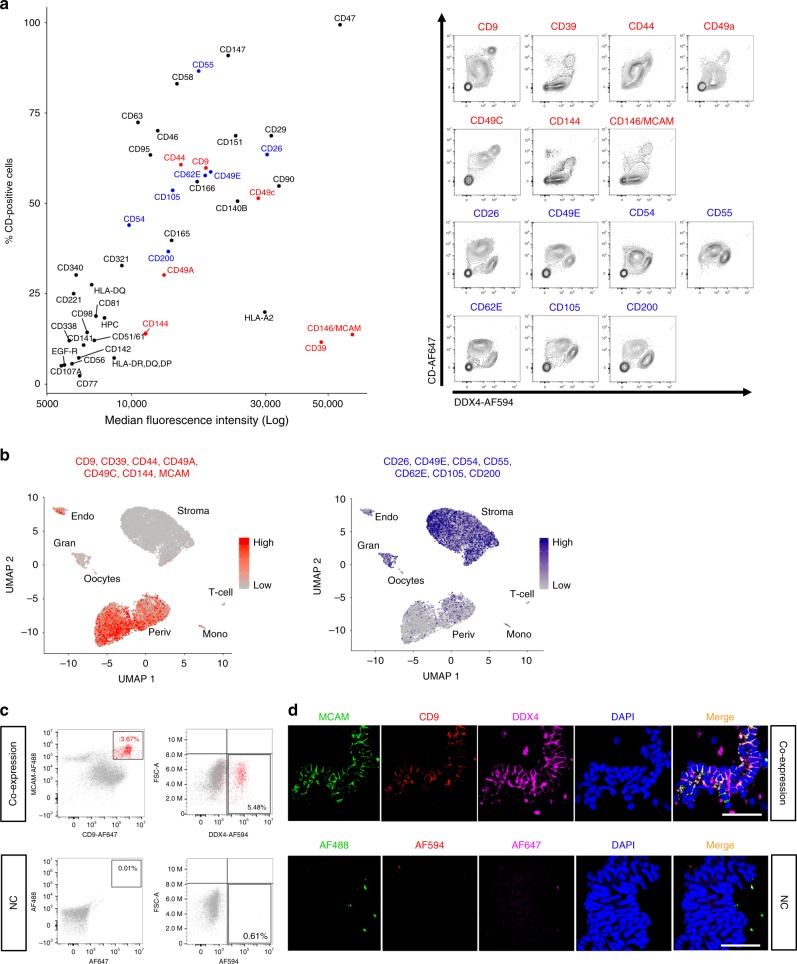
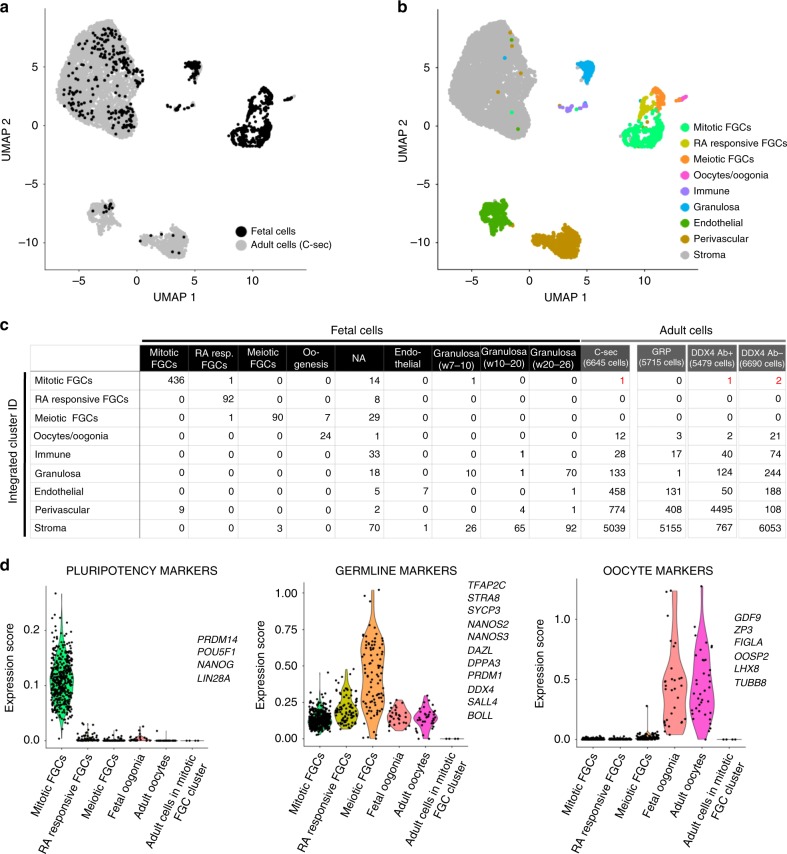
The human ovary orchestrates sex hormone production and undergoes monthly structural changes to release mature oocytes. The outer lining of the ovary (cortex) has a key role in defining fertility in women as it harbors the ovarian reserve. It has been postulated that putative oogonial stem cells exist in the ovarian cortex and that these can be captured by DDX4 antibody isolation. Here, we report single-cell transcriptomes and cell surface antigen profiles of over 24,000 cells from high quality ovarian cortex samples from 21 patients. Our data identify transcriptional profiles of six main cell types; oocytes, granulosa cells, immune cells, endothelial cells, perivascular cells, and stromal cells. Cells captured by DDX4 antibody are perivascular cells, not oogonial stem cells. Our data do not support the existence of germline stem cells in adult human ovaries, thereby reinforcing the dogma of a limited ovarian reserve.
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
Ovary does harbor stem cells - size of the cells matter!J Ovarian Res. 2020 Apr 17;13(1):39. doi: 10.1186/s13048-020-00647-2. J Ovarian Res. 2020. PMID: 32303227 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
