

Patient-derived small intestinal myofibroblasts direct perfused, physiologically responsive capillary development in a microfluidic Gut-on-a-Chip Model
- PMID: 32123209
- PMCID: PMC7051952
- DOI: 10.1038/s41598-020-60672-5
Patient-derived small intestinal myofibroblasts direct perfused, physiologically responsive capillary development in a microfluidic Gut-on-a-Chip Model
Abstract
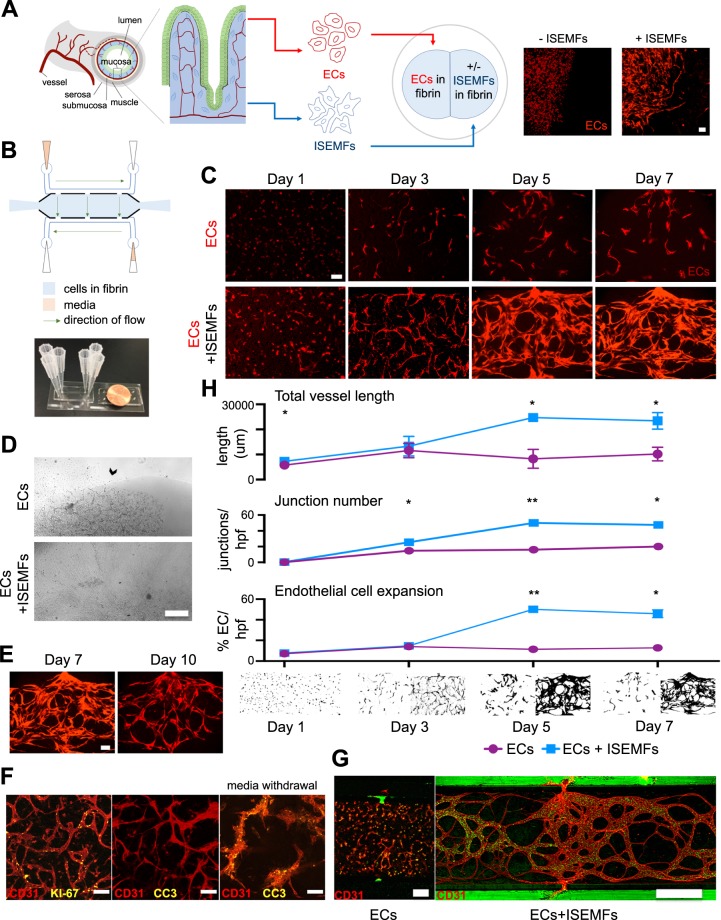
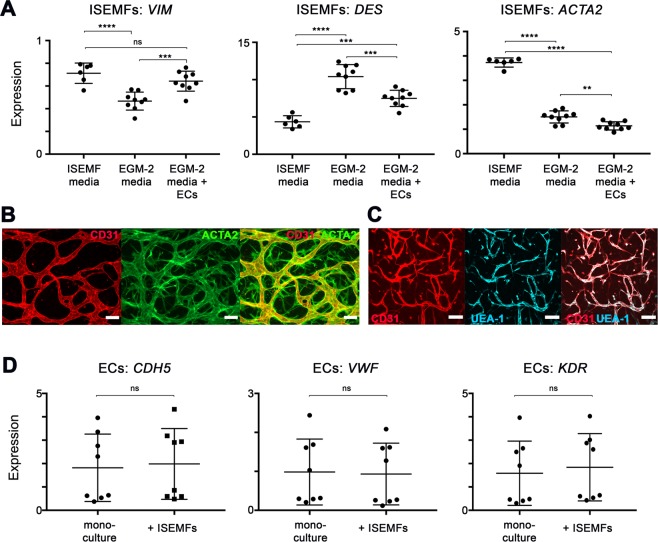
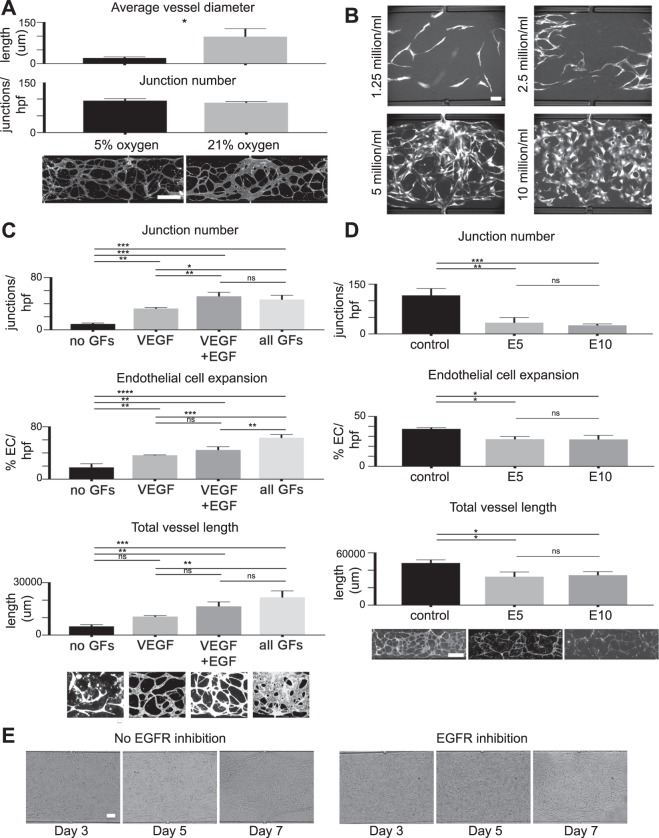
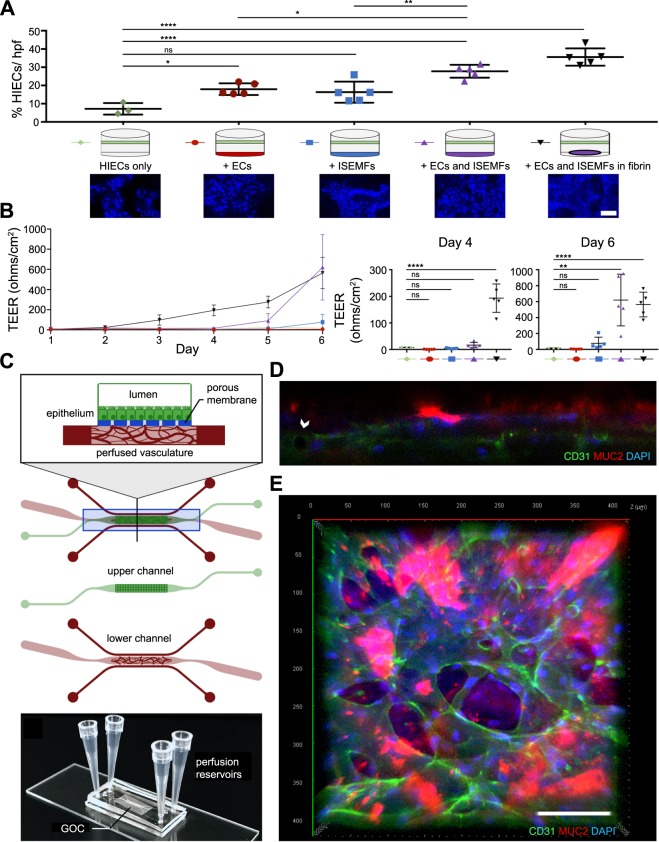
The development and physiologic role of small intestine (SI) vasculature is poorly studied. This is partly due to a lack of targetable, organ-specific markers for in vivo studies of two critical tissue components: endothelium and stroma. This challenge is exacerbated by limitations of traditional cell culture techniques, which fail to recapitulate mechanobiologic stimuli known to affect vessel development. Here, we construct and characterize a 3D in vitro microfluidic model that supports the growth of patient-derived intestinal subepithelial myofibroblasts (ISEMFs) and endothelial cells (ECs) into perfused capillary networks. We report how ISEMF and EC-derived vasculature responds to physiologic parameters such as oxygen tension, cell density, growth factors, and pharmacotherapy with an antineoplastic agent (Erlotinib). Finally, we demonstrate effects of ISEMF and EC co-culture on patient-derived human intestinal epithelial cells (HIECs), and incorporate perfused vasculature into a gut-on-a-chip (GOC) model that includes HIECs. Overall, we demonstrate that ISEMFs possess angiogenic properties as evidenced by their ability to reliably, reproducibly, and quantifiably facilitate development of perfused vasculature in a microfluidic system. We furthermore demonstrate the feasibility of including perfused vasculature, including ISEMFs, as critical components of a novel, patient-derived, GOC system with translational relevance as a platform for precision and personalized medicine research.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Epimorphin regulates the intestinal stem cell niche via effects on the stromal microenvironment.Am J Physiol Gastrointest Liver Physiol. 2018 Aug 1;315(2):G185-G194. doi: 10.1152/ajpgi.00224.2017. Epub 2018 Apr 6. Am J Physiol Gastrointest Liver Physiol. 2018. PMID: 29631377 Free PMC article.
-
Intestinal subepithelial myofibroblasts support the growth of intestinal epithelial stem cells.PLoS One. 2014 Jan 6;9(1):e84651. doi: 10.1371/journal.pone.0084651. eCollection 2014. PLoS One. 2014. PMID: 24400106 Free PMC article.
-
Intestinal subepithelial myofibroblasts support in vitro and in vivo growth of human small intestinal epithelium.PLoS One. 2011;6(11):e26898. doi: 10.1371/journal.pone.0026898. Epub 2011 Nov 17. PLoS One. 2011. PMID: 22125602 Free PMC article.
-
Intestinal Models for Personalized Medicine: from Conventional Models to Microfluidic Primary Intestine-on-a-chip.Stem Cell Rev Rep. 2022 Aug;18(6):2137-2151. doi: 10.1007/s12015-021-10205-y. Epub 2021 Jun 28. Stem Cell Rev Rep. 2022. PMID: 34181185 Free PMC article. Review.
-
Organ-on-Chip Approaches for Intestinal 3D In Vitro Modeling.Cell Mol Gastroenterol Hepatol. 2022;13(2):351-367. doi: 10.1016/j.jcmgh.2021.08.015. Epub 2021 Aug 25. Cell Mol Gastroenterol Hepatol. 2022. PMID: 34454168 Free PMC article. Review.
Cited by
-
Advances in the Model Structure of In Vitro Vascularized Organ-on-a-Chip.Cyborg Bionic Syst. 2024 Apr 25;5:0107. doi: 10.34133/cbsystems.0107. eCollection 2024. Cyborg Bionic Syst. 2024. PMID: 40353137 Free PMC article.
-
Prognostic value of RNA methylation-related genes in gastric adenocarcinoma based on bioinformatics.PeerJ. 2024 Feb 29;12:e16951. doi: 10.7717/peerj.16951. eCollection 2024. PeerJ. 2024. PMID: 38436027 Free PMC article.
-
Salivary Gland Tissue Engineering Approaches: State of the Art and Future Directions.Cells. 2021 Jul 8;10(7):1723. doi: 10.3390/cells10071723. Cells. 2021. PMID: 34359893 Free PMC article. Review.
-
Gut-on-a-Chip Models: Current and Future Perspectives for Host-Microbial Interactions Research.Biomedicines. 2023 Feb 18;11(2):619. doi: 10.3390/biomedicines11020619. Biomedicines. 2023. PMID: 36831155 Free PMC article. Review.
-
Organ-on-a-Chip systems for new drugs development.ADMET DMPK. 2021 Mar 22;9(2):111-141. doi: 10.5599/admet.942. eCollection 2021. ADMET DMPK. 2021. PMID: 35299767 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
- R01 HD105301/HD/NICHD NIH HHS/United States
- R01 DK106382/DK/NIDDK NIH HHS/United States
- T32 DK007130/DK/NIDDK NIH HHS/United States
- R01 DK104698/DK/NIDDK NIH HHS/United States
- R01 DK114047/DK/NIDDK NIH HHS/United States
- R03 DK111473/DK/NIDDK NIH HHS/United States
- R01 DK109384/DK/NIDDK NIH HHS/United States
- R01 DK118568/DK/NIDDK NIH HHS/United States
- R01 DK112378/DK/NIDDK NIH HHS/United States
- K08 DK101608/DK/NIDDK NIH HHS/United States
- P30 DK052574/DK/NIDDK NIH HHS/United States
- T32 HD043010/HD/NICHD NIH HHS/United States
- K01 DK109081/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Research Materials
