

Tau Reduction Prevents Key Features of Autism in Mouse Models
- PMID: 32126198
- PMCID: PMC7210056
- DOI: 10.1016/j.neuron.2020.01.038
Tau Reduction Prevents Key Features of Autism in Mouse Models
Abstract
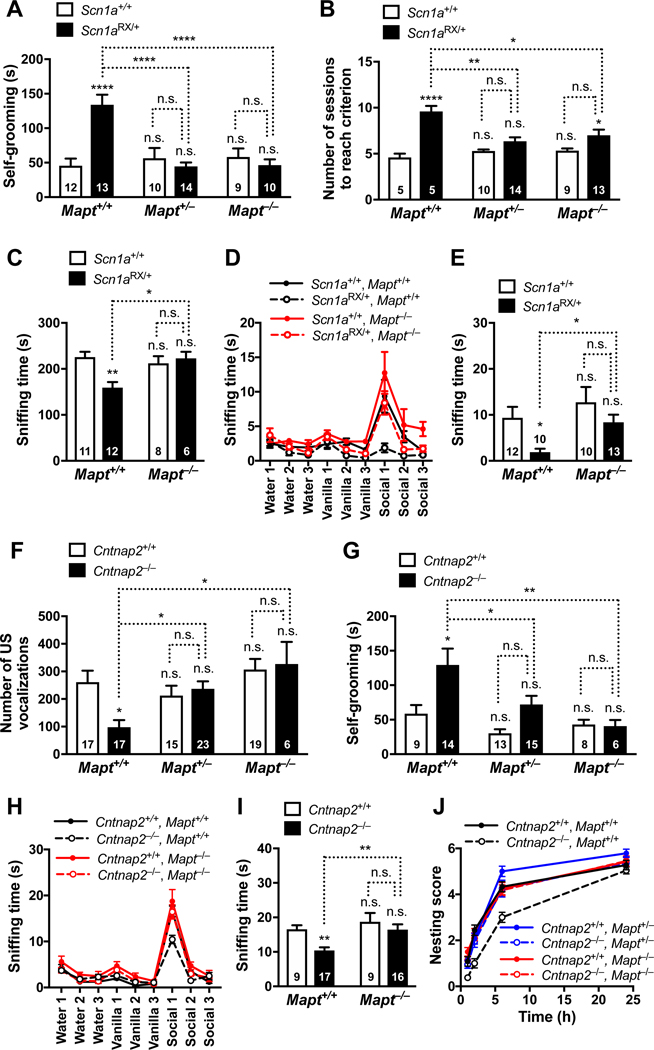
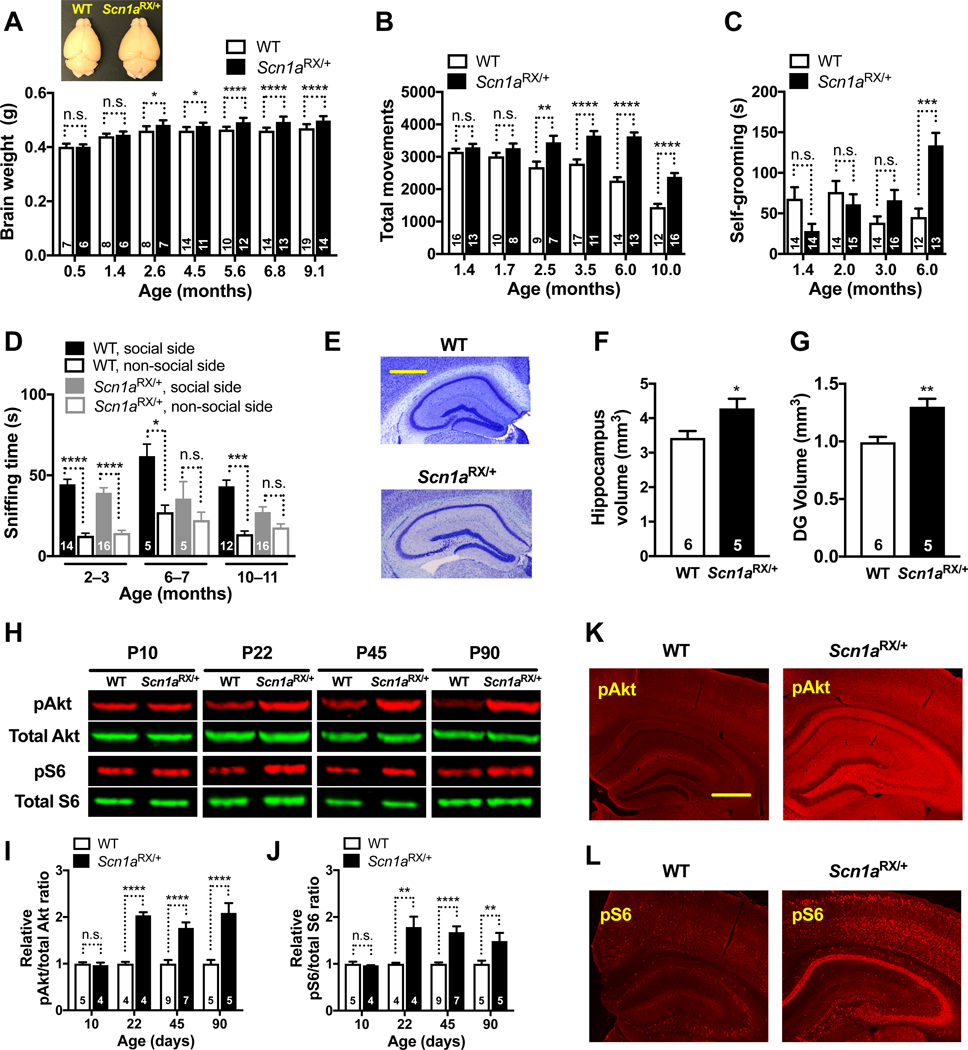
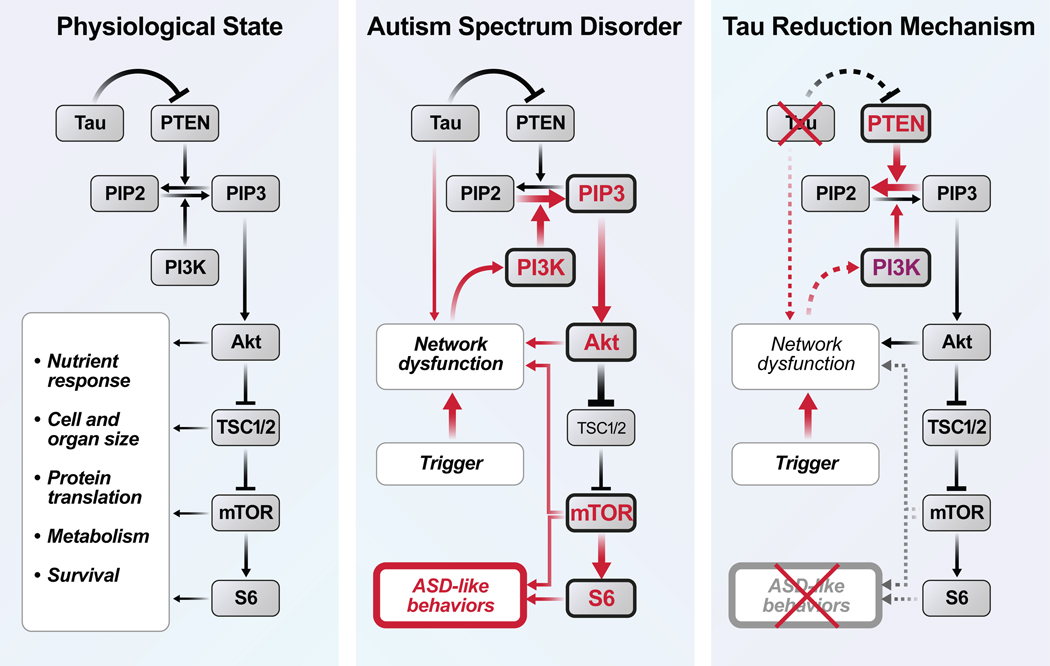
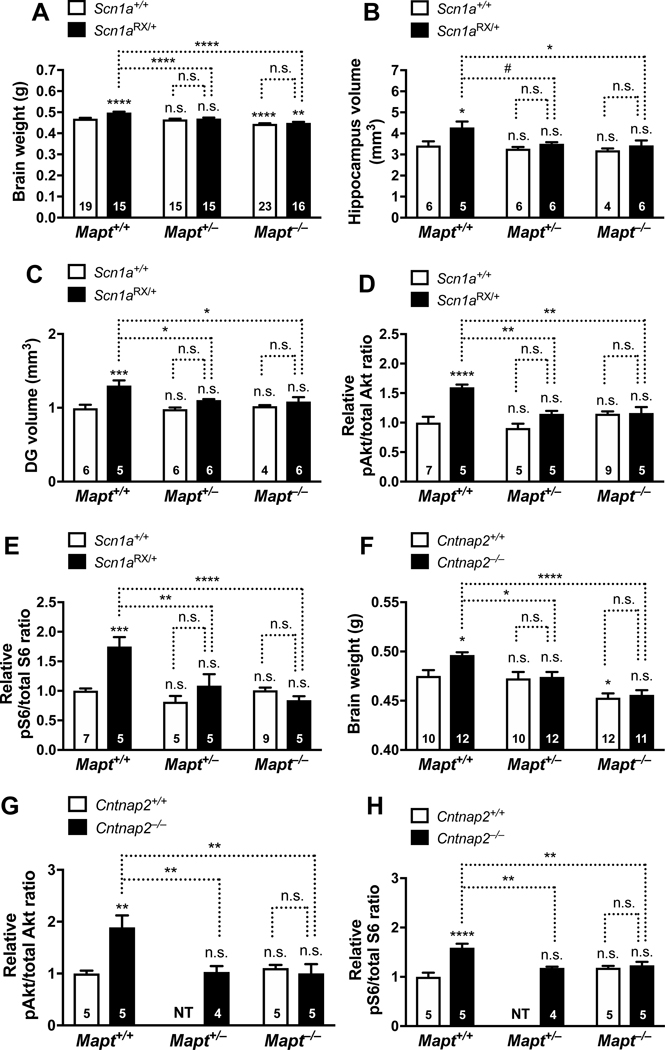
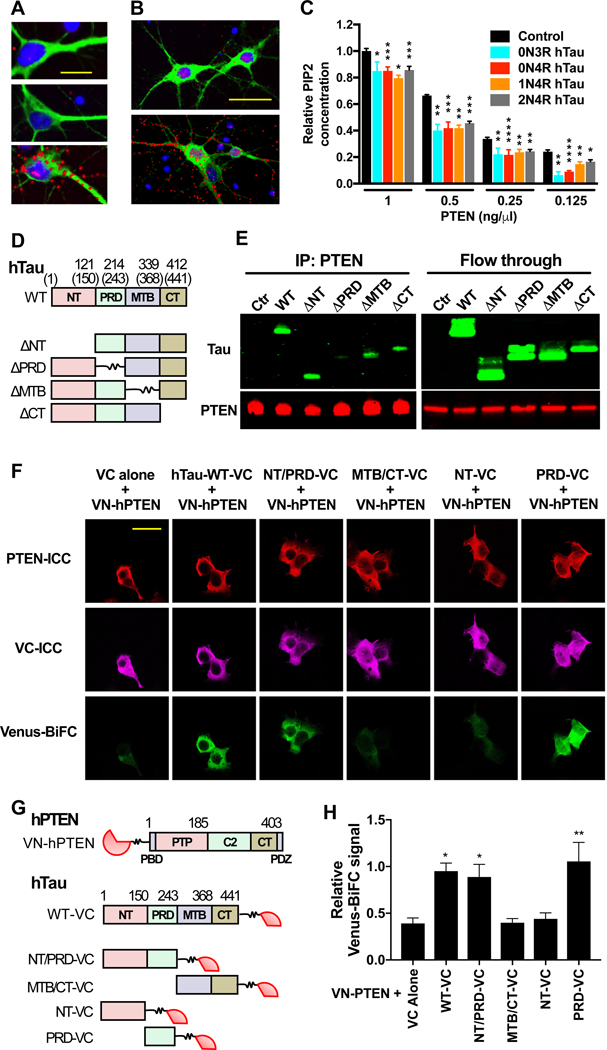
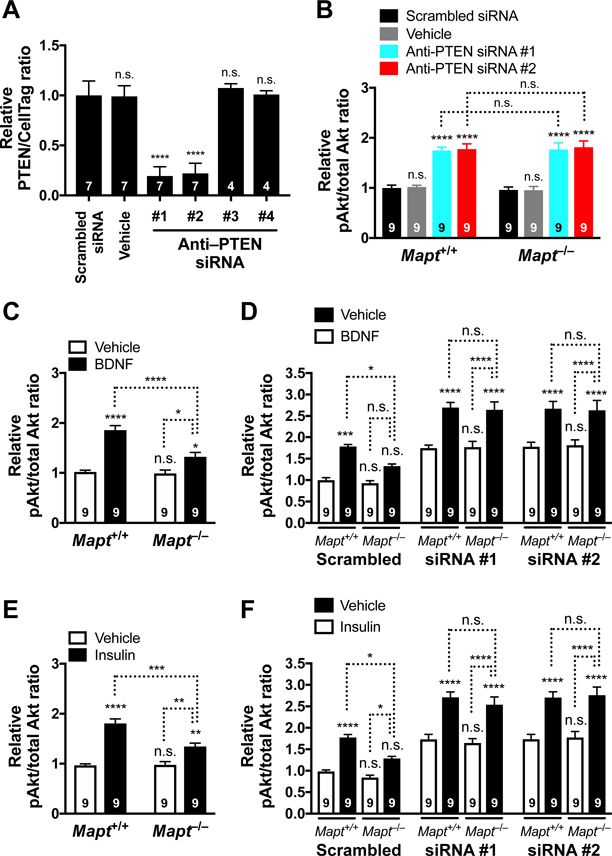
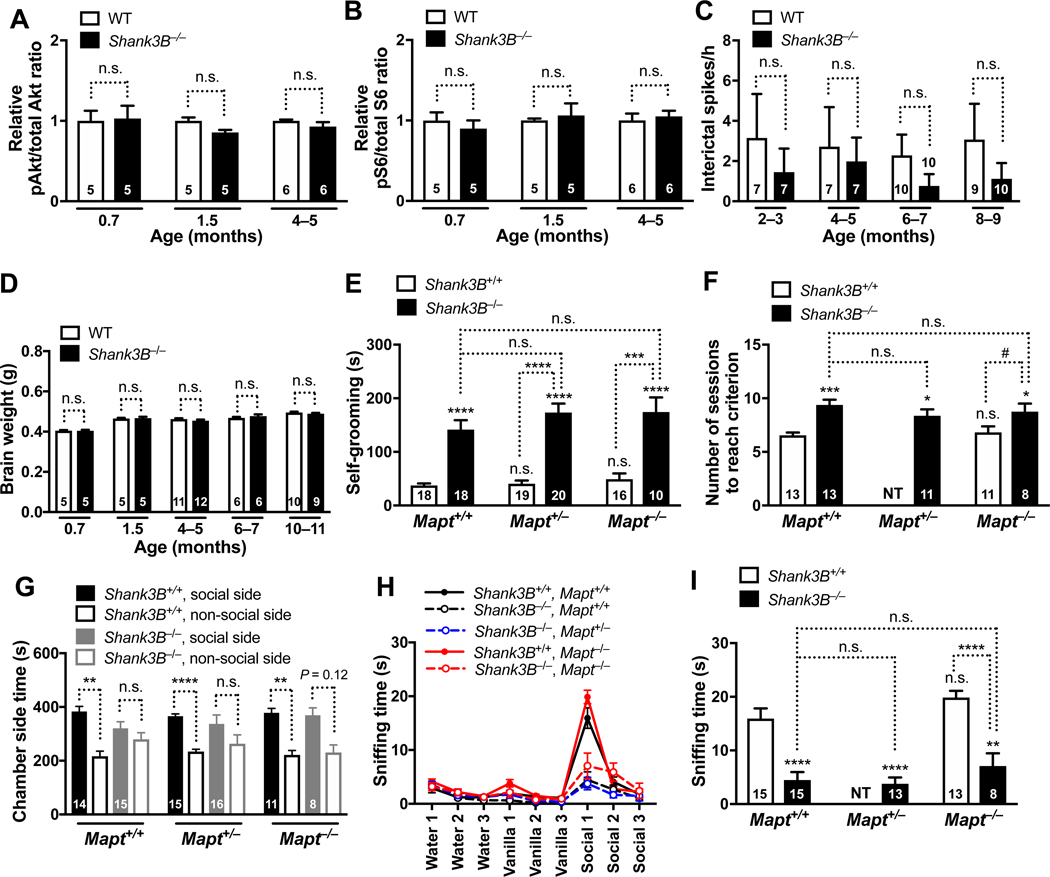
Autism is characterized by repetitive behaviors, impaired social interactions, and communication deficits. It is a prevalent neurodevelopmental disorder, and available treatments offer little benefit. Here, we show that genetically reducing the protein tau prevents behavioral signs of autism in two mouse models simulating distinct causes of this condition. Similar to a proportion of people with autism, both models have epilepsy, abnormally enlarged brains, and overactivation of the phosphatidylinositol 3-kinase (PI3K)/Akt (protein kinase B)/ mammalian target of rapamycin (mTOR) signaling pathway. All of these abnormalities were prevented or markedly diminished by partial or complete genetic removal of tau. We identify disinhibition of phosphatase and tensin homolog deleted on chromosome 10 (PTEN), a negative PI3K regulator that tau controls, as a plausible mechanism and demonstrate that tau interacts with PTEN via tau's proline-rich domain. Our findings suggest an enabling role of tau in the pathogenesis of autism and identify tau reduction as a potential therapeutic strategy for some of the disorders that cause this condition.
Keywords: Akt; Cntnap2; PI3 kinase; PTEN; Scn1a; Shank3; autism spectrum disorders; mTOR; megalencephaly; tau.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests L.M. is a co-inventor on patents filed by the Gladstone Institutes that focus on tau reduction as a strategy to block neural network dysfunction (U.S. patent number 9,189,982 for “Agents that Reduce Neuronal Overexcitation,” issued December 1, 2015; U.S. patent number 9,084,813 for “Agents that Reduce Neuronal Overexcitation,” issued July 21, 2015). He is the principal investigator on a research project to develop tau-lowering small-molecule drugs that is supported, in part, by a corporate-sponsored research agreement between the Gladstone Institutes and Cure Network Dolby Acceleration Partners. He also serves on the scientific advisory boards of Arvinas Operations, Biogen, and Dolby Family Ventures and has provided consulting services to Sangamo Therapeutics.
Figures
Comment in
-
Tau: A Novel Entry Point for mTOR-Based Treatments in Autism Spectrum Disorder?Neuron. 2020 May 6;106(3):359-361. doi: 10.1016/j.neuron.2020.04.019. Neuron. 2020. PMID: 32380047
References
-
- Allen Institute for Brain Science. (2019). Allen Brain Atlas: Developing Mouse Brain. Available from: https://developingmouse.brain-map.org/gene/show/17529.
-
- Baio J., Wiggins L., Christensen DL., Maenner MJ., Daniels J., Warren Z., Kurzius-Spencer M., Zahorodny W., Robinson Rosenberg C., White T., et al. (2018). Prevalence of autism spectrum disorder among children aged 8 years—Autism and Developmental Disabilities Monitoring Network, 11 Sites, United States, 2014. MMWR Surveill Summ 67, 1–23. - PMC - PubMed
-
- Berkvens JJ, Veugen I, Veendrick-Meekes MJ, Snoeijen-Schouwenaars FM, Schelhaas HJ, Willemsen MH, Tan IY, and Aldenkamp AP (2015). Autism and behavior in adult patients with Dravet syndrome (DS). Epilepsy Behav 47, 11–16. - PubMed
-
- Bidinosti M, Botta P, Kruttner S, Proenca CC, Stoehr N, Bernhard M, Fruh I, Mueller M, Bonenfant D, Voshol H, et al. (2016). CLK2 inhibition ameliorates autistic features associated with SHANK3 deficiency. Science 351, 1199–1203. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
