

Sensory neuron-derived NaV1.7 contributes to dorsal horn neuron excitability
- PMID: 32128393
- PMCID: PMC7030926
- DOI: 10.1126/sciadv.aax4568
Sensory neuron-derived NaV1.7 contributes to dorsal horn neuron excitability
Abstract
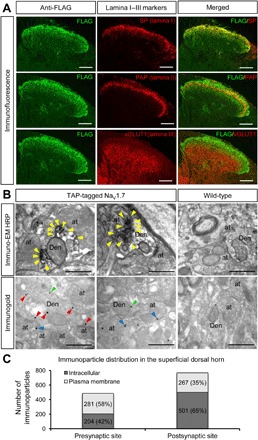
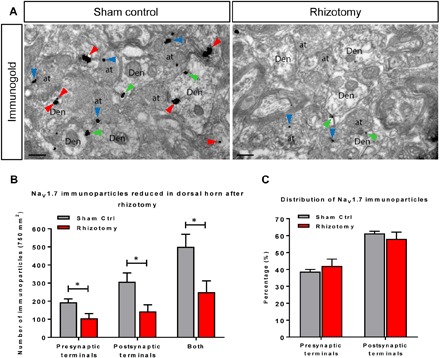
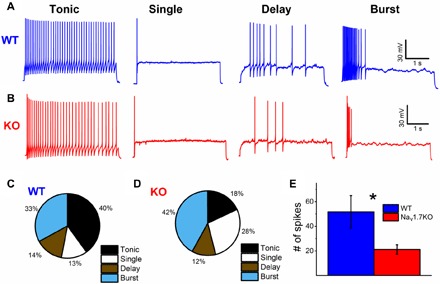
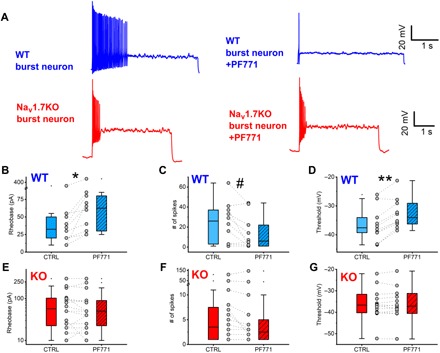
Expression of the voltage-gated sodium channel NaV1.7 in sensory neurons is required for pain sensation. We examined the role of NaV1.7 in the dorsal horn of the spinal cord using an epitope-tagged NaV1.7 knock-in mouse. Immuno-electron microscopy showed the presence of NaV1.7 in dendrites of superficial dorsal horn neurons, despite the absence of mRNA. Rhizotomy of L5 afferent nerves lowered the levels of NaV1.7 in the dorsal horn. Peripheral nervous system-specific NaV1.7 null mutant mice showed central deficits, with lamina II dorsal horn tonic firing neurons more than halved and single spiking neurons more than doubled. NaV1.7 blocker PF05089771 diminished excitability in dorsal horn neurons but had no effect on NaV1.7 null mutant mice. These data demonstrate an unsuspected functional role of primary afferent neuron-generated NaV1.7 in dorsal horn neurons and an expression pattern that would not be predicted by transcriptomic analysis.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution License 4.0 (CC BY).
Figures
References
-
- Habib A. M., Matsuyama A., Okorokov A. L., Santana-Varela S., Bras J. T., Aloisi A. M., Emery E. C., Bogdanov Y. D., Follenfant M., Gossage S. J., Gras M., Humphrey J., Kolesnikov A., Le Cann K., Li S., Minett M. S., Pereira V., Ponsolles C., Sikandar S., Torres J. M., Yamaoka K., Zhao J., Komine Y., Yamamori T., Maniatis N., Panov K. I., Houlden H., Ramirez J. D., Bennett D. L. H., Marsili L., Bachiocco V., Wood J. N., Cox J. J., A novel human pain insensitivity disorder caused by a point mutation in ZFHX2. Brain 141, 365–376 (2018). - PMC - PubMed
-
- Cox J. J., Reimann F., Nicholas A. K., Thornton G., Roberts E., Springell K., Karbani G., Jafri H., Mannan J., Raashid Y., al-Gazali L., Hamamy H., Valente E. M., Gorman S., Williams R., McHale D. P., Wood J. N., Gribble F. M., Woods C. G., An SCN9A channelopathy causes congenital inability to experience pain. Nature 444, 894–898 (2006). - PMC - PubMed
-
- Sexton J. E., Cox J. J., Zhao J., Wood J. N., The genetics of pain: Implications for therapeutics. Annu. Rev. Pharmacol. Toxicol. 58, 123–142 (2018). - PubMed
-
- Habib A. M., Wood J. N., Cox J. J., Sodium channels and pain. Handb. Exp. Pharmacol. 227, 39–56 (2015). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
