

Long noncoding RNA metastasis-associated lung adenocarcinoma transcript 1 cooperates with enhancer of zeste homolog 2 to promote hepatocellular carcinoma development by modulating the microRNA-22/Snail family transcriptional repressor 1 axis
- PMID: 32129914
- PMCID: PMC7226208
- DOI: 10.1111/cas.14372
Long noncoding RNA metastasis-associated lung adenocarcinoma transcript 1 cooperates with enhancer of zeste homolog 2 to promote hepatocellular carcinoma development by modulating the microRNA-22/Snail family transcriptional repressor 1 axis
Abstract
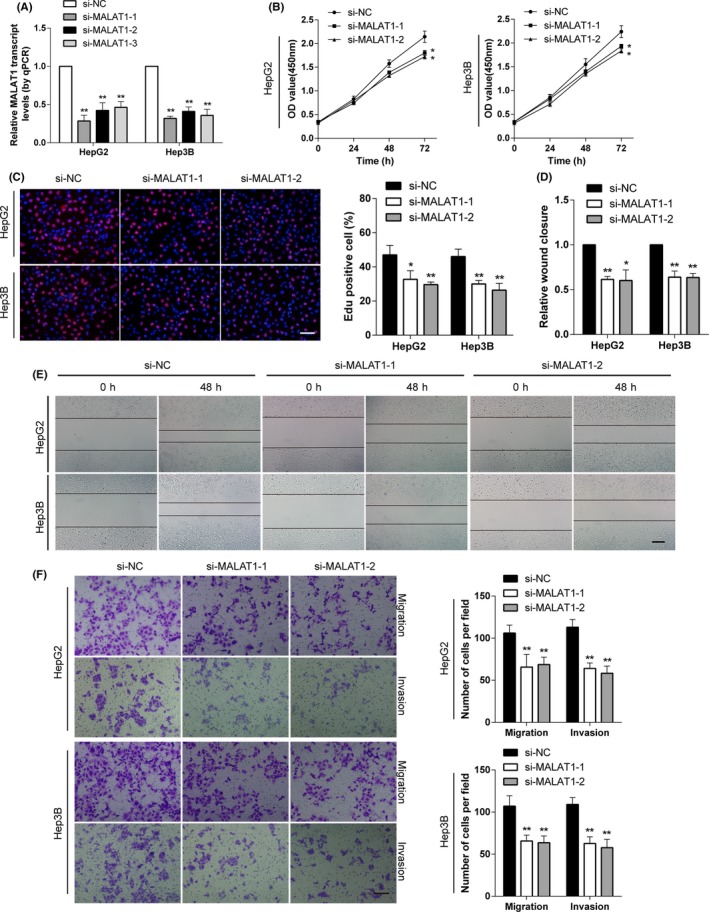
Metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) is an oncogenic long noncoding RNA that has been found to promote carcinogenesis and metastasis in many tumors. However, the underlying role of MALAT1 in the progression and metastasis of hepatocellular carcinoma (HCC) remains unclear. In this study, aberrantly elevated levels of MALAT1 were detected in both HCC specimens and cell lines. We found that knockdown of MALAT1 caused retardation in proliferation, migration, and invasion both in vivo and in vitro. Mechanistic investigations showed that Snail family transcriptional repressor 1 (SNAI1) is a direct target of microRNA (miR)-22 and that MALAT1 modulates SNAI1 expression by acting as a competing endogenous RNA for miR-22. Inhibition of miR-22 restored SNAI1 expression suppressed by MALAT1 knockdown. Furthermore, MALAT1 facilitated the enrichment of enhancer of zeste homolog 2 (EZH2) at the promoter region of miR-22 and E-cadherin, which was repressed by MALAT1 knockdown. Cooperating with EZH2, MALAT1 positively regulated SNAI1 by repressing miR-22 and inhibiting E-cadherin expression, playing a vital role in epithelial to mesenchymal transition. In conclusion, our results reveal a mechanism by which MALAT1 promotes HCC progression and provides a potential target for HCC therapy.
Keywords: EZH2; MALAT1; SNAI1; hepatocellular carcinoma; miR-22.
© 2020 The Authors. Cancer Science published by John Wiley & Sons Australia, Ltd on behalf of Japanese Cancer Association.
Conflict of interest statement
The authors have no conflict of interest for this article.
Figures






References
-
- El‐Serag HB, Rudolph KL. Hepatocellular carcinoma: epidemiology and molecular carcinogenesis. Gastroenterology. 2007;132:2557‐2576. - PubMed
-
- Li H, Wu K, Tao K, et al. Tim‐3/galectin‐9 signaling pathway mediates T‐cell dysfunction and predicts poor prognosis in patients with hepatitis B virus‐associated hepatocellular carcinoma. Hepatology. 2012;56:1342‐1351. - PubMed
-
- Chen W, Zheng R, Baade PD, et al. Cancer statistics in China, 2015. CA Cancer J Clin. 2016;66:115‐132. - PubMed
-
- Siegel RL, Miller KD, Jemal A. Cancer statistics, 2016. CA Cancer J Clin. 2016;66(1):7–30. - PubMed
MeSH terms
Substances
Grants and funding
- 2019CFB496/Natural Science Foundation of Hubei Province
- 2017KFYXJJ251/Huazhong University of Science and Technology Innovation Research Fund
- 2015CB5540007/National Key Basic Research Program of China
- 81772967/National Natural Science Foundation of China
- 2017KFYXJJ251/Funds for training young and middle-aged medical backbone talents in Wuhan
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
