

IL-21 is a broad negative regulator of IgE class switch recombination in mouse and human B cells
- PMID: 32130409
- PMCID: PMC7201927
- DOI: 10.1084/jem.20190472
IL-21 is a broad negative regulator of IgE class switch recombination in mouse and human B cells
Abstract
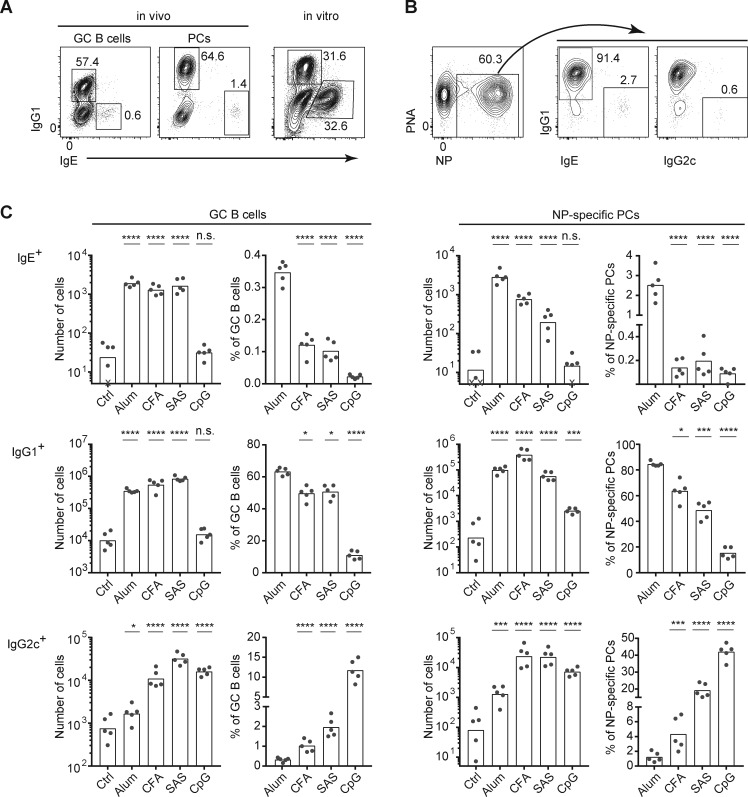
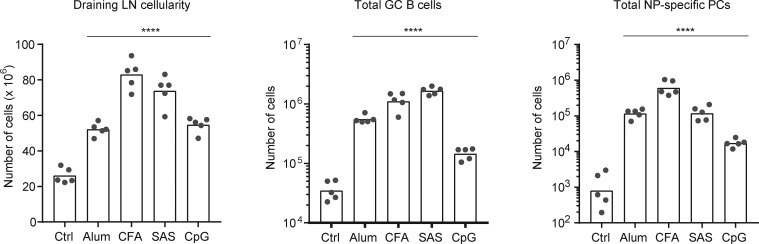
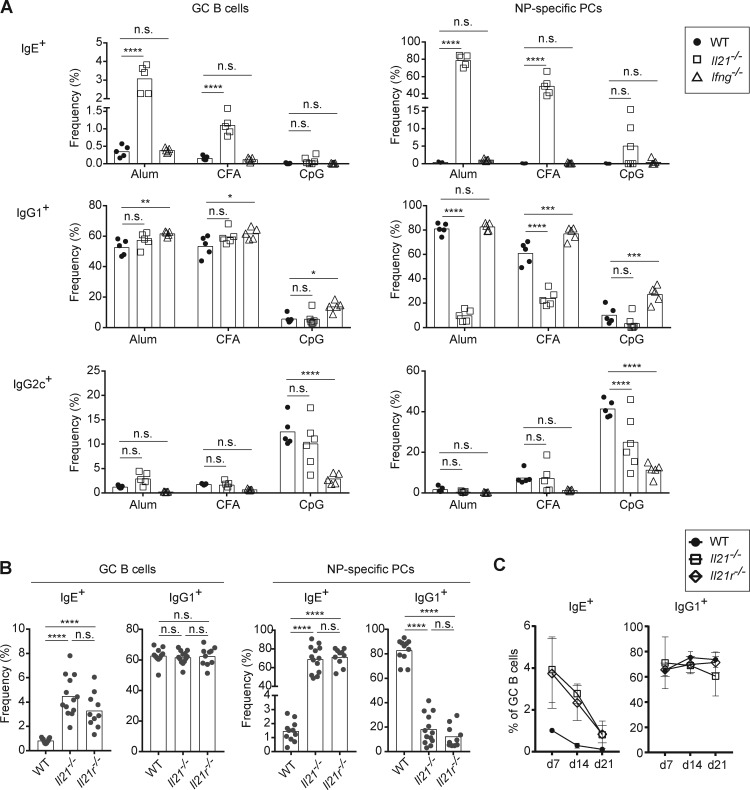
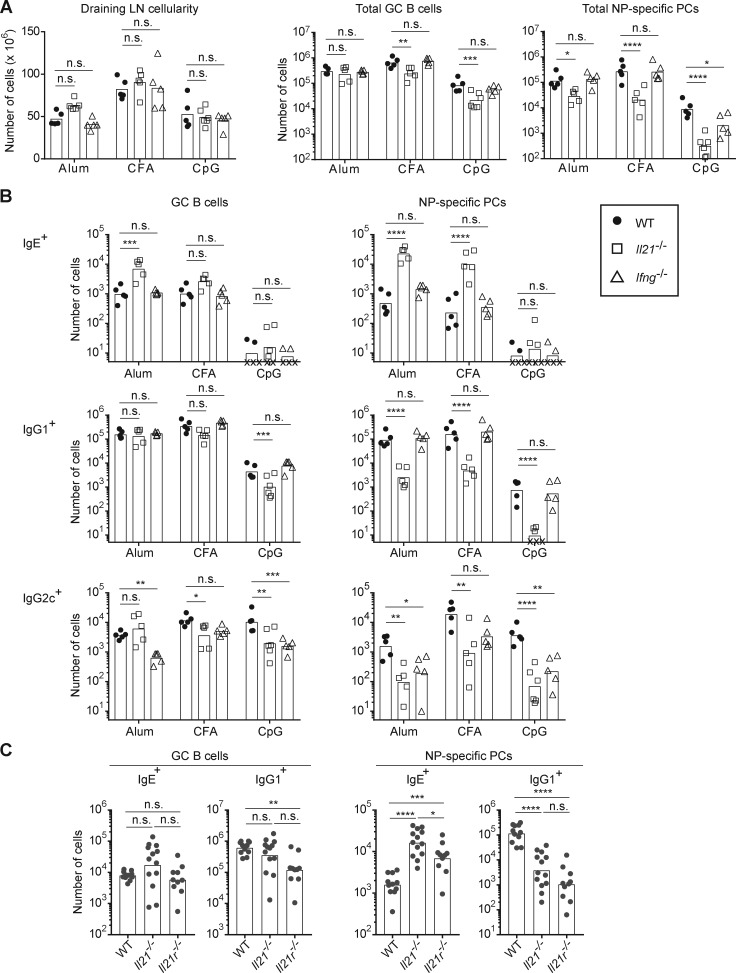
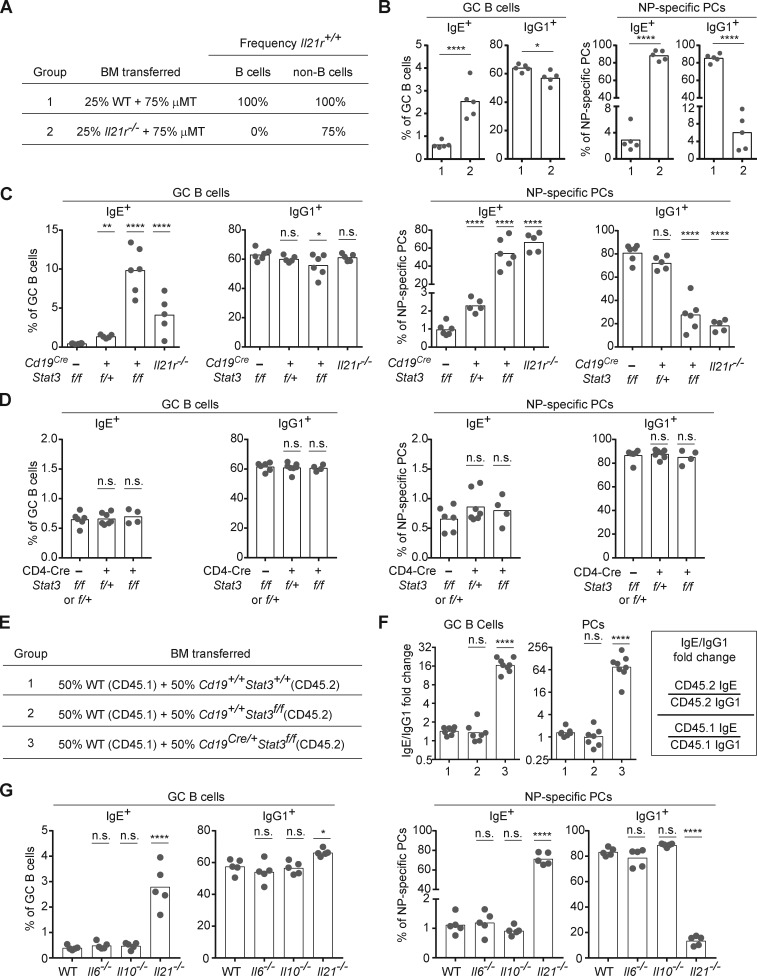
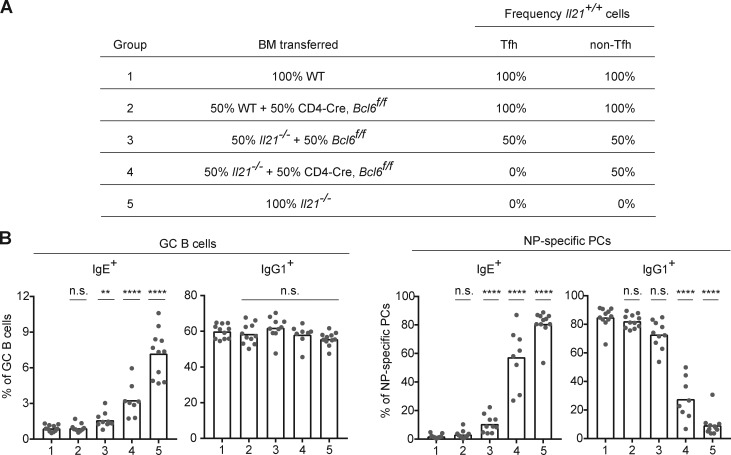
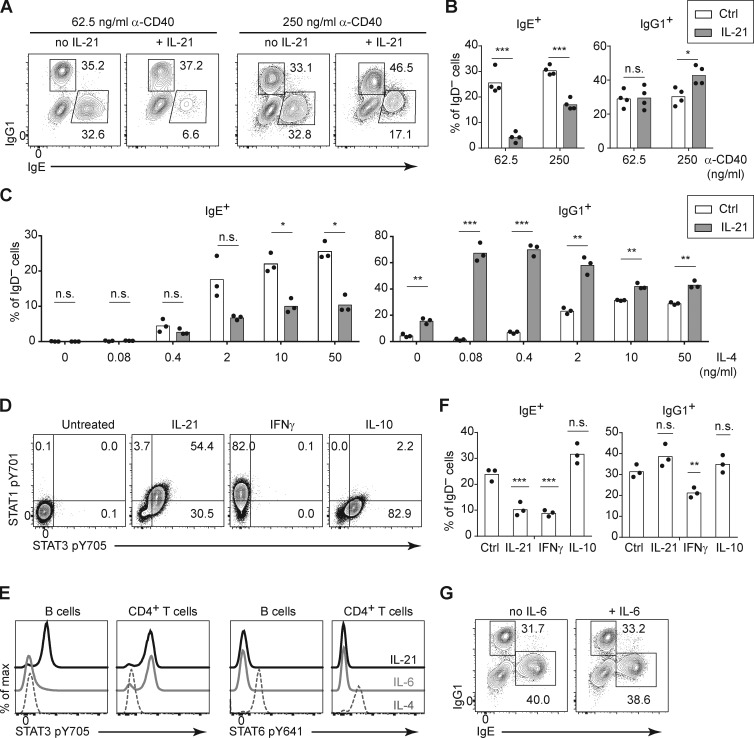
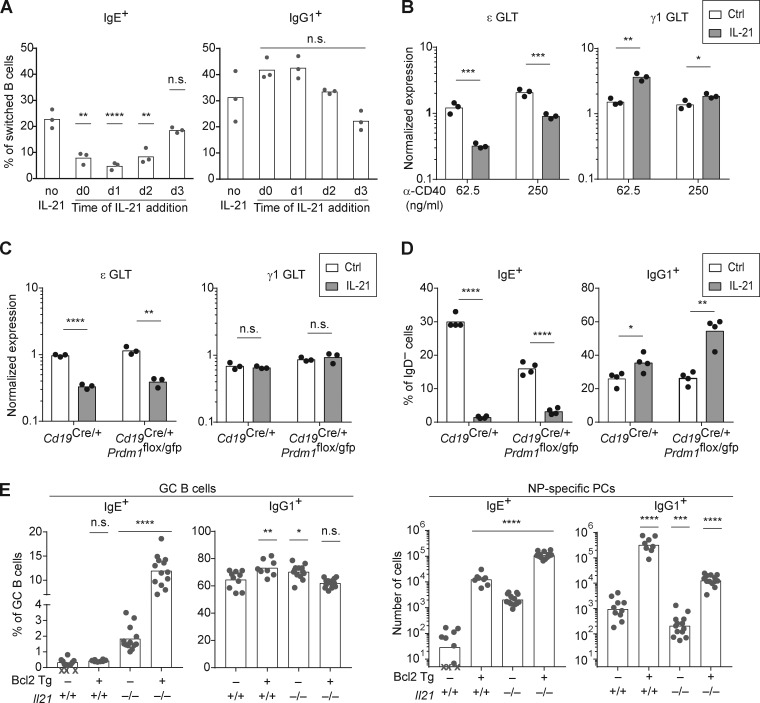
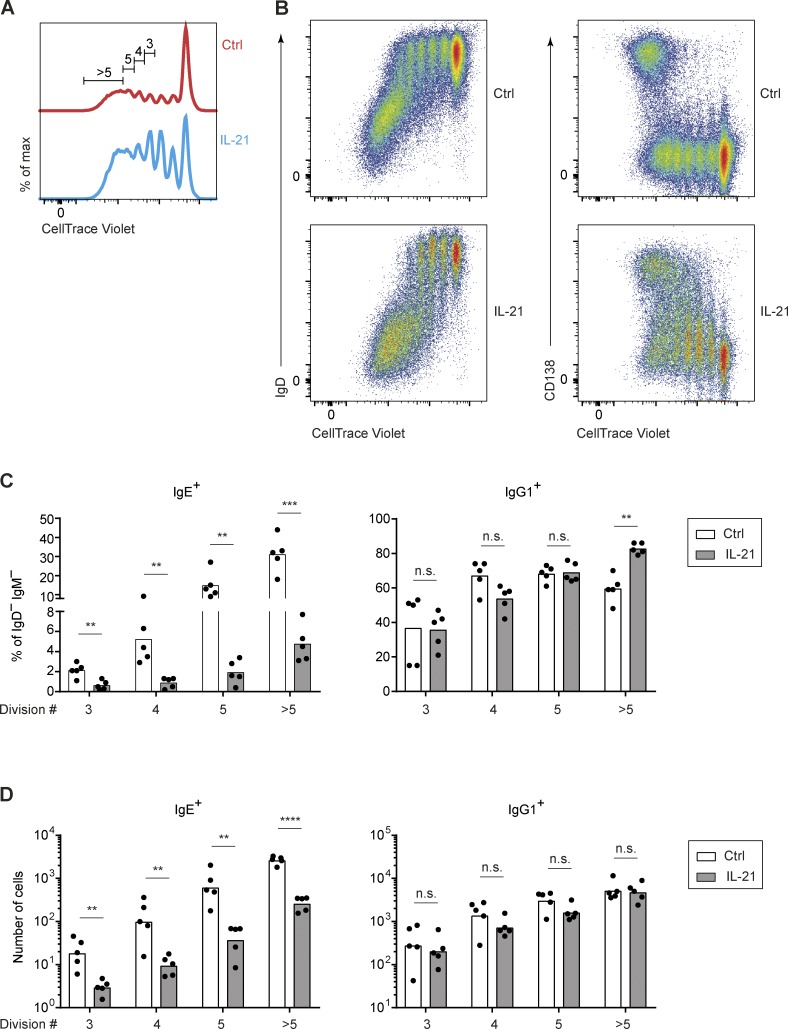
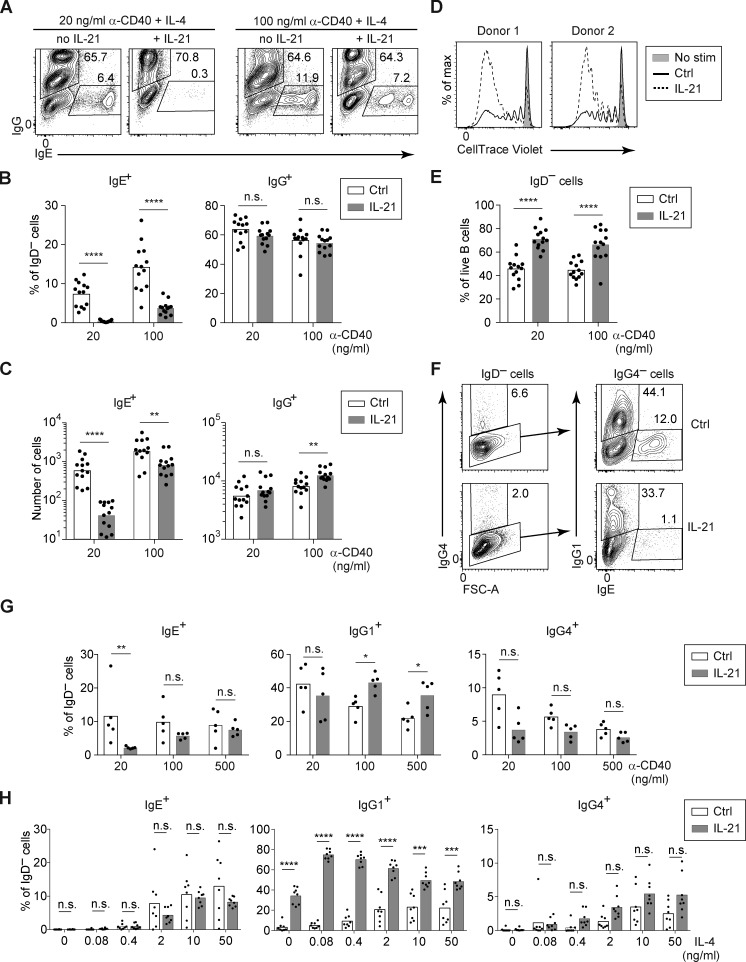
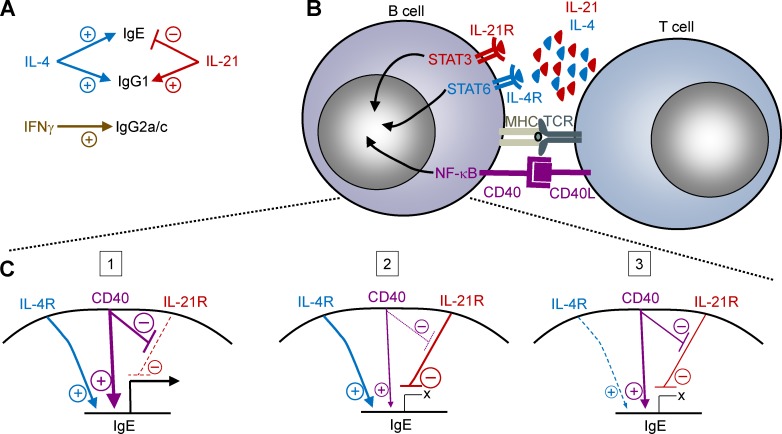
IgE antibodies may elicit potent allergic reactions, and their production is tightly controlled. The tendency to generate IgE has been thought to reflect the balance between type 1 and type 2 cytokines, with the latter promoting IgE. Here, we reevaluated this paradigm by a direct cellular analysis, demonstrating that IgE production was not limited to type 2 immune responses yet was generally constrained in vivo. IL-21 was a critical negative regulator of IgE responses, whereas IFN-γ, IL-6, and IL-10 were dispensable. Follicular helper T cells were the primary source of IL-21 that inhibited IgE responses by directly engaging the IL-21 receptor on B cells and triggering STAT3-dependent signaling. We reconciled previous discordant results between mouse and human B cells and revealed that the inhibition of IgE class switch recombination by IL-21 was attenuated by CD40 signaling, whereas IgG1 class switch recombination was potentiated by IL-21 in the context of limited IL-4. These findings establish key features of the extrinsic regulation of IgE production by cytokines.
© 2020 Yang et al.
Conflict of interest statement
Disclosures: The authors declare no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
