

Lactation stage impacts the glycolytic function of bovine CD4+ T cells during ex vivo activation
- PMID: 32132555
- PMCID: PMC7055328
- DOI: 10.1038/s41598-020-60691-2
Lactation stage impacts the glycolytic function of bovine CD4+ T cells during ex vivo activation
Abstract
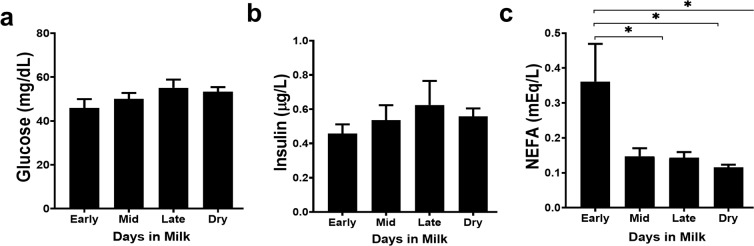
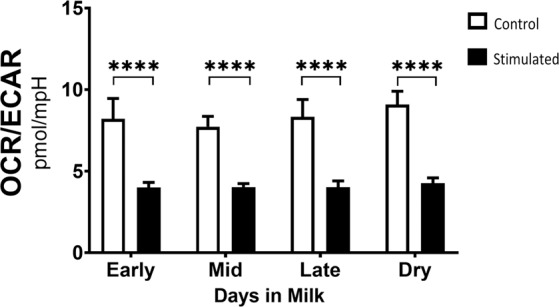
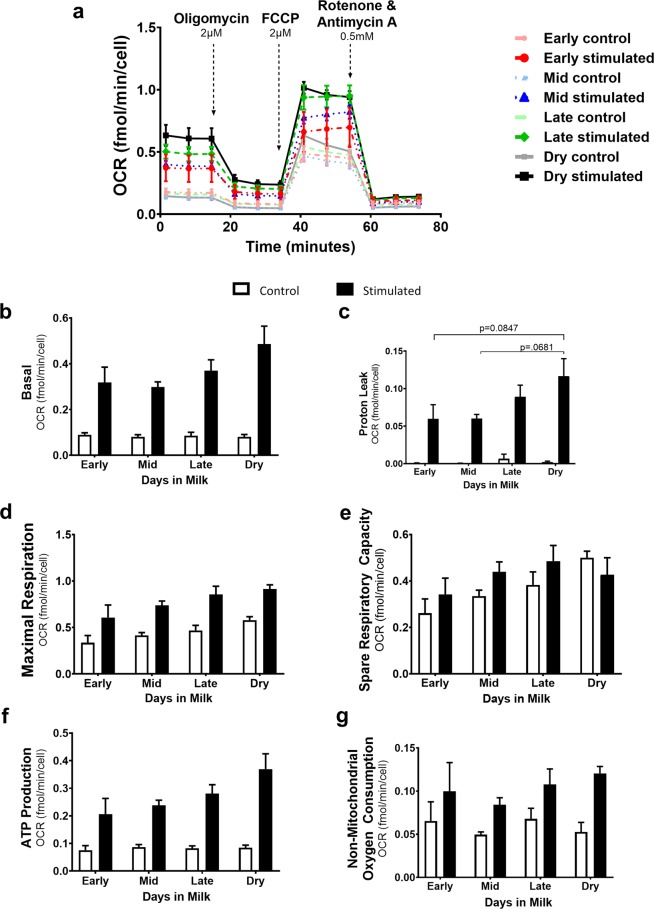
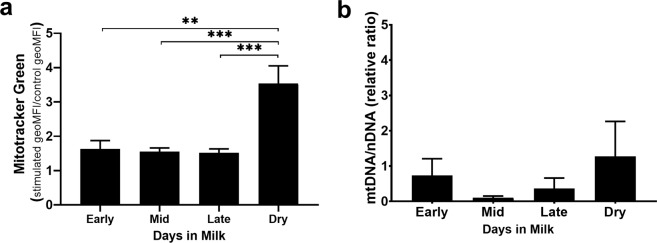
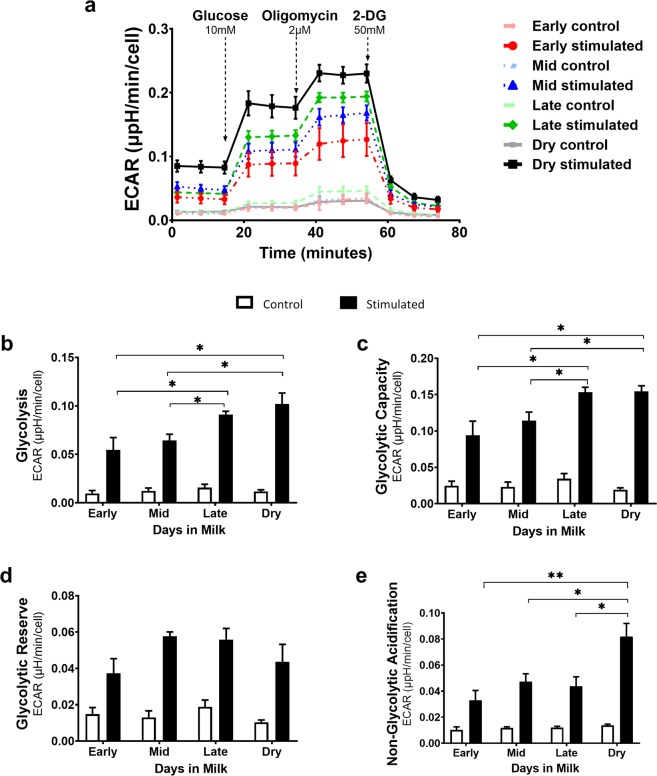
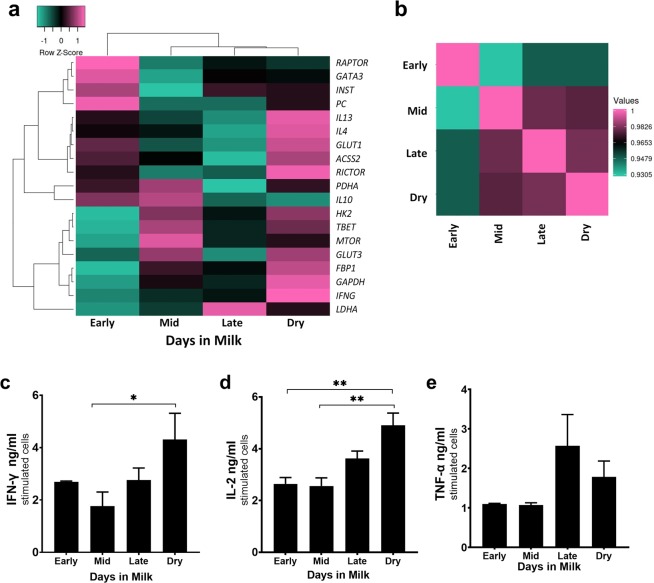
Dairy cattle undergo dynamic physiological changes over the course of a full lactation into the dry period, which impacts their immunocompetence. During activation, T cells undergo a characteristic rewiring to increase the uptake of glucose and metabolically reprogram to favor aerobic glycolysis over oxidative phosphorylation. To date it remains to be completely elucidated how the altered energetic demands associated with lactation in dairy cows impacts T cell metabolic reprogramming. Thus, in our ex vivo studies we have examined the influence of stage of lactation (early lactation into the dry period) on cellular metabolism in activated bovine CD4+ T cells. Results showed higher rates of glycolytic function in activated CD4+ T cells from late lactation and dry cows compared to cells from early and mid-lactation cows. Similarly, protein and mRNA expression of cytokines were higher in CD4+ T cells from dry cows than CD4+ T cells from lactating cows. The data suggest CD4+ T cells from lactating cows have an altered metabolic responsiveness that could impact the immunocompetence of these animals, particularly those in early lactation, and increase their susceptibility to infection.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Ingvartsen KL, Moyes KM. Factors contributing to immunosuppression in the dairy cow during the periparturient period. Jpn. J. Vet. Res. 2015;63(Suppl 1):S15–24. - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
