

Comparative Analysis of CPI-Motif Regulation of Biochemical Functions of Actin Capping Protein
- PMID: 32133840
- PMCID: PMC7301643
- DOI: 10.1021/acs.biochem.0c00092
Comparative Analysis of CPI-Motif Regulation of Biochemical Functions of Actin Capping Protein
Abstract
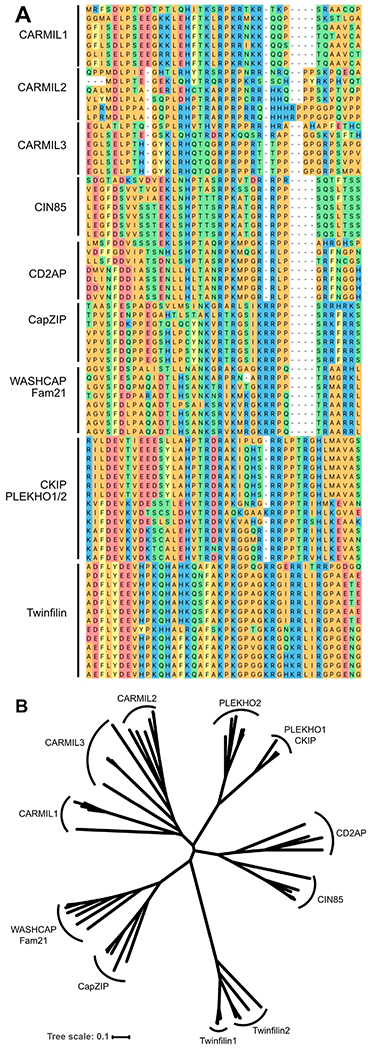
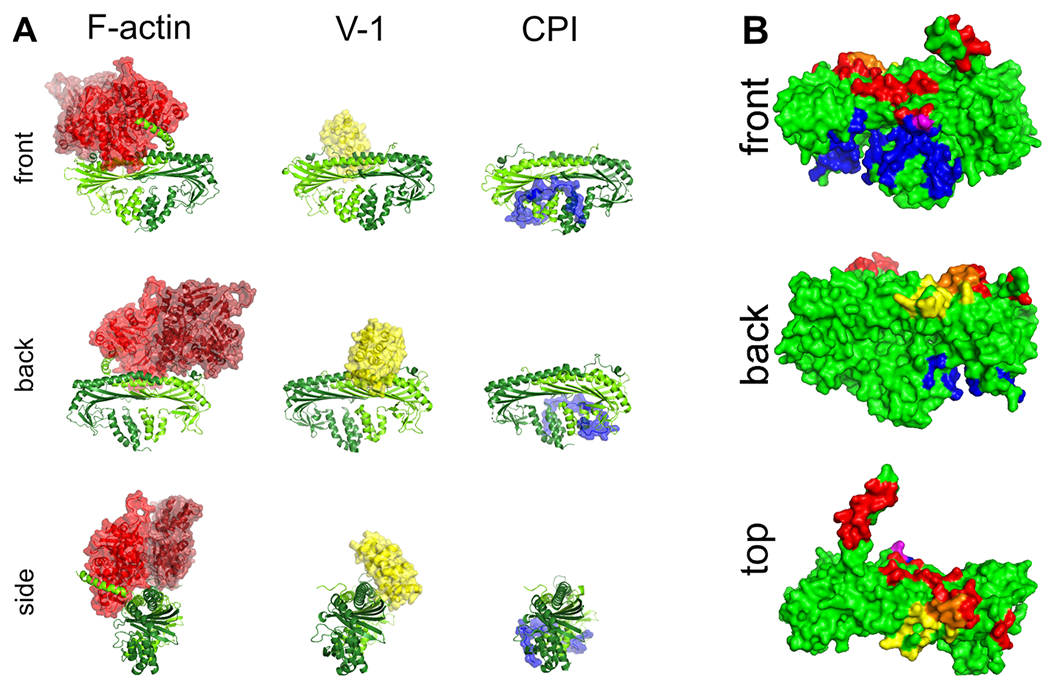
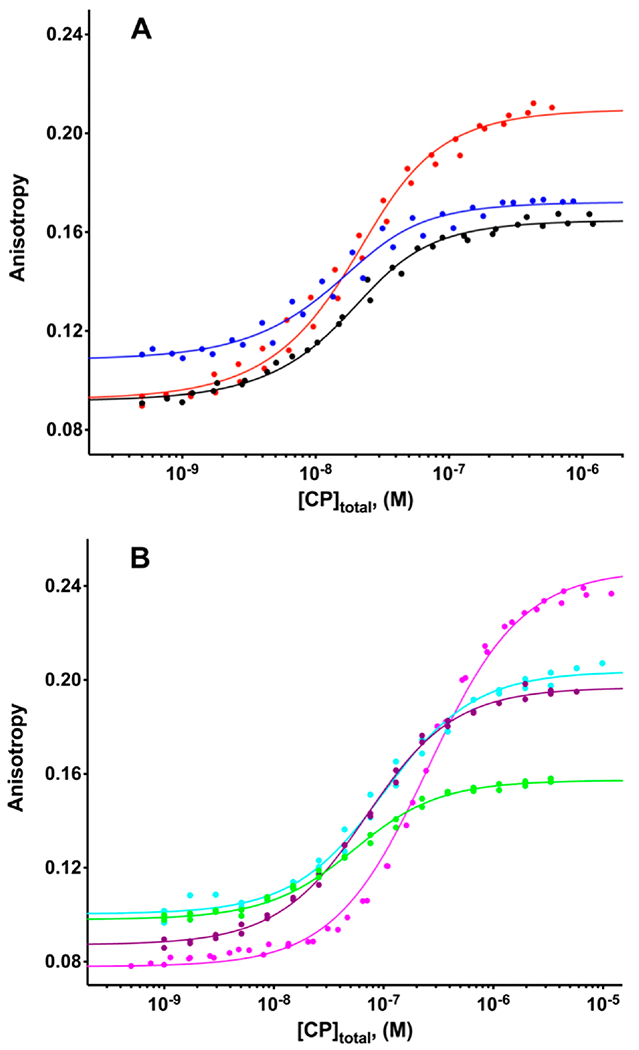
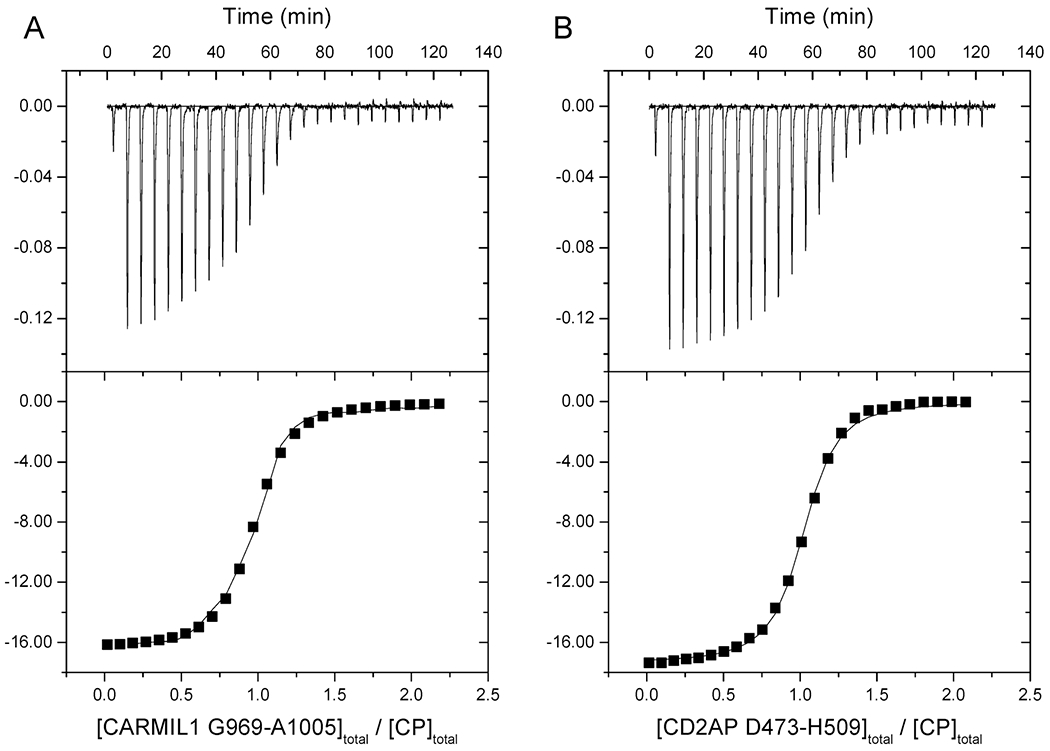
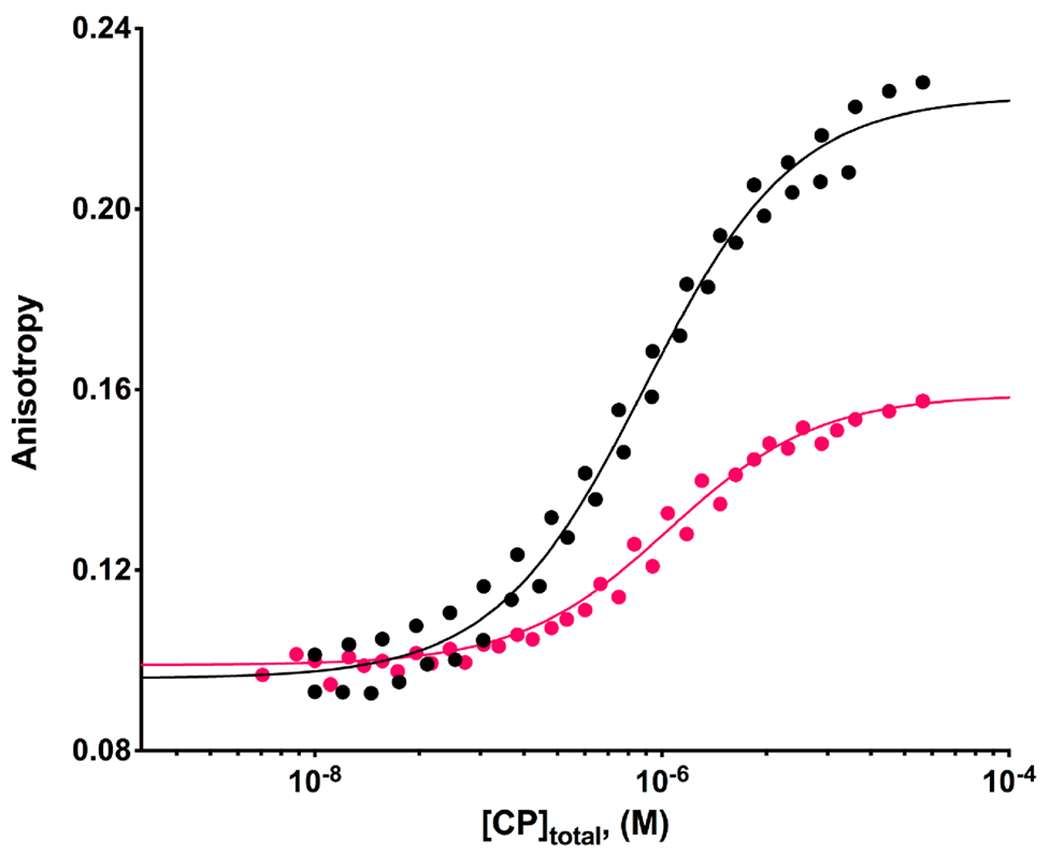
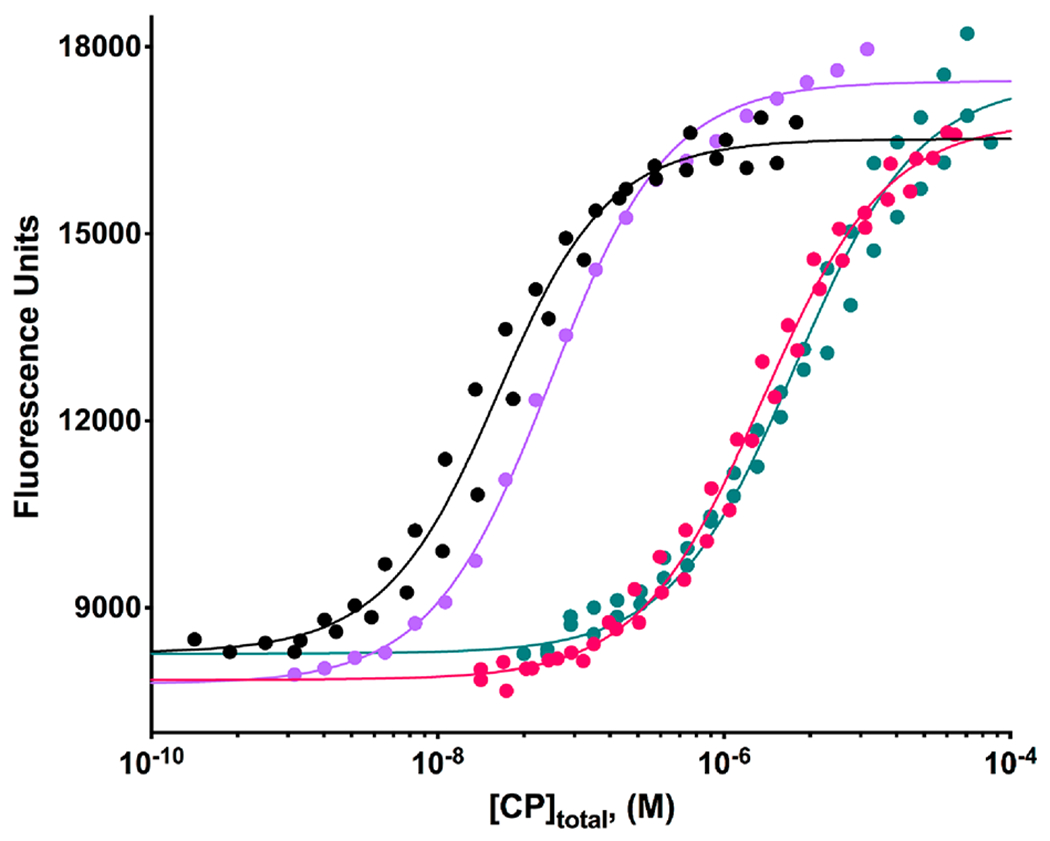
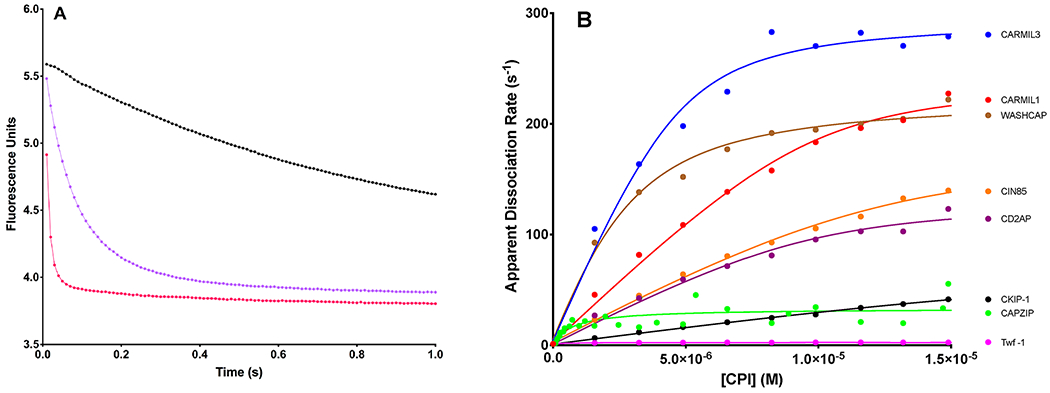
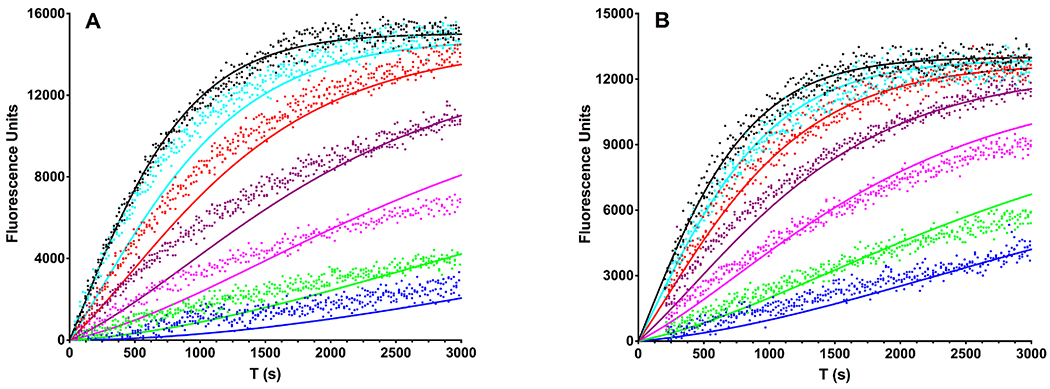
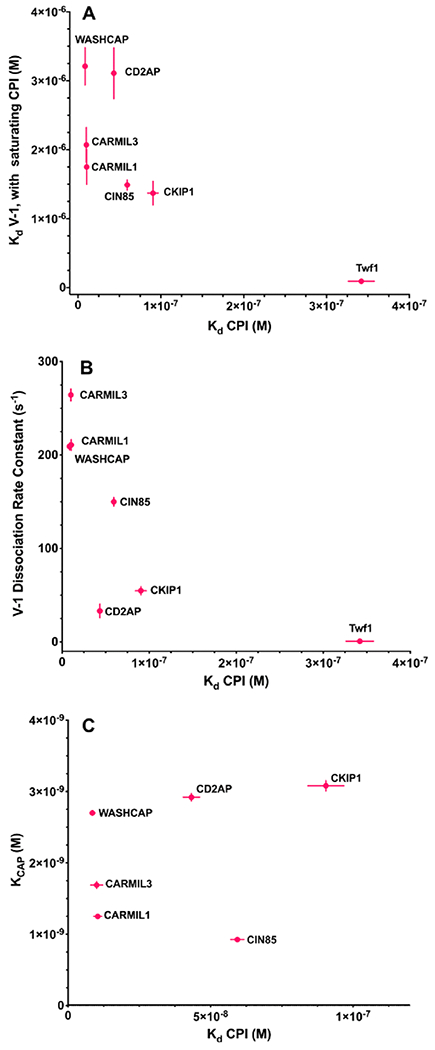
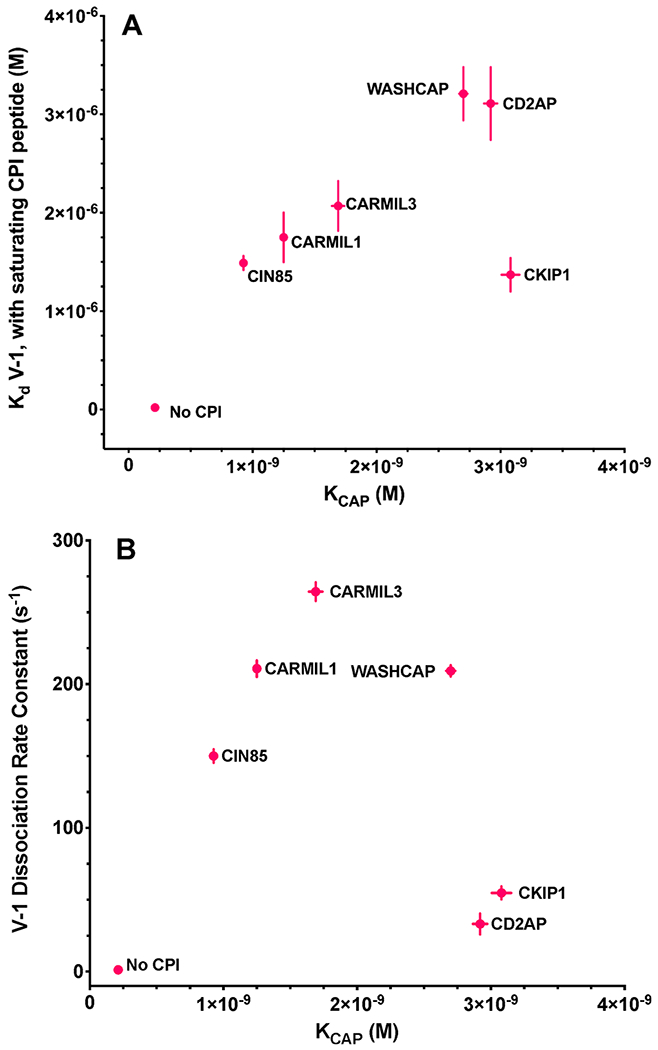
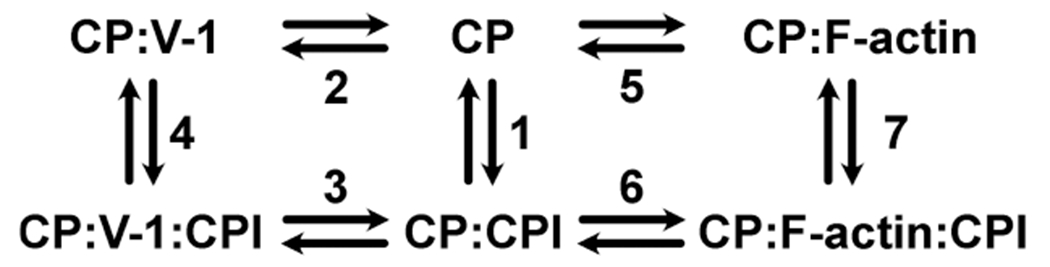
The heterodimeric actin capping protein (CP) is regulated by a set of proteins that contain CP-interacting (CPI) motifs. Outside of the CPI motif, the sequences of these proteins are unrelated and distinct. The CPI motif and surrounding sequences are conserved within a given protein family, when compared to those of other CPI-motif protein families. Using biochemical assays with purified proteins, we compared the ability of CPI-motif-containing peptides from different protein families (a) to bind to CP, (b) to allosterically inhibit barbed-end capping by CP, and (c) to allosterically inhibit interaction of CP with V-1, another regulator of CP. We found large differences in potency among the different CPI-motif-containing peptides, and the different functional assays showed different orders of potency. These biochemical differences among the CPI-motif peptides presumably reflect interactions between CP and CPI-motif peptides involving amino acid residues that are conserved but are not part of the strictly defined consensus, as it was originally identified in comparisons of sequences of CPI motifs across all protein families [Hernandez-Valladares, M., et al. (2010) Structural characterization of a capping protein interaction motif defines a family of actin filament regulators. Nat. Struct. Mol. Biol. 17, 497-503; Bruck, S., et al. (2006) Identification of a Novel Inhibitory Actin-capping Protein Binding Motif in CD2-associated Protein. J. Biol. Chem. 281, 19196-19203]. These biochemical differences may be important for conserved distinct functions of CPI-motif protein families in cells with respect to the regulation of CP activity and actin assembly near membranes.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
