

Topological structure and dynamics of three-dimensional active nematics
- PMID: 32139540
- PMCID: PMC7984424
- DOI: 10.1126/science.aaz4547
Topological structure and dynamics of three-dimensional active nematics
Abstract
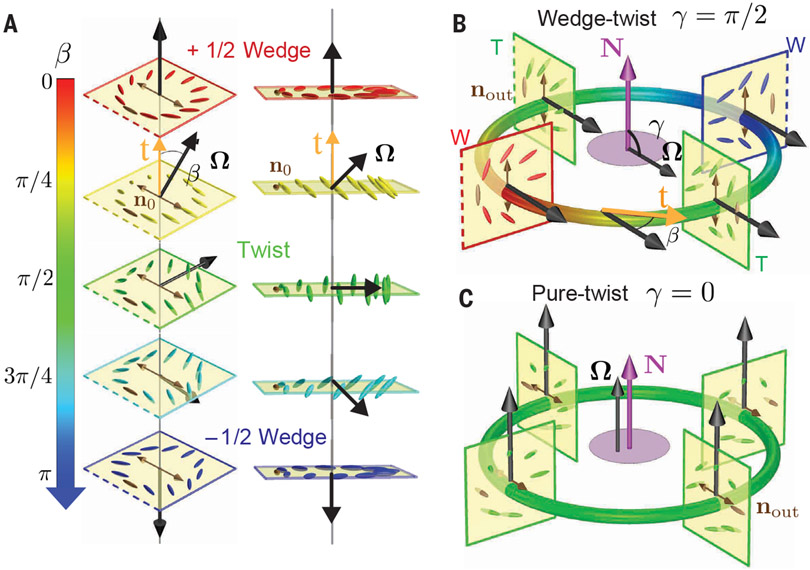
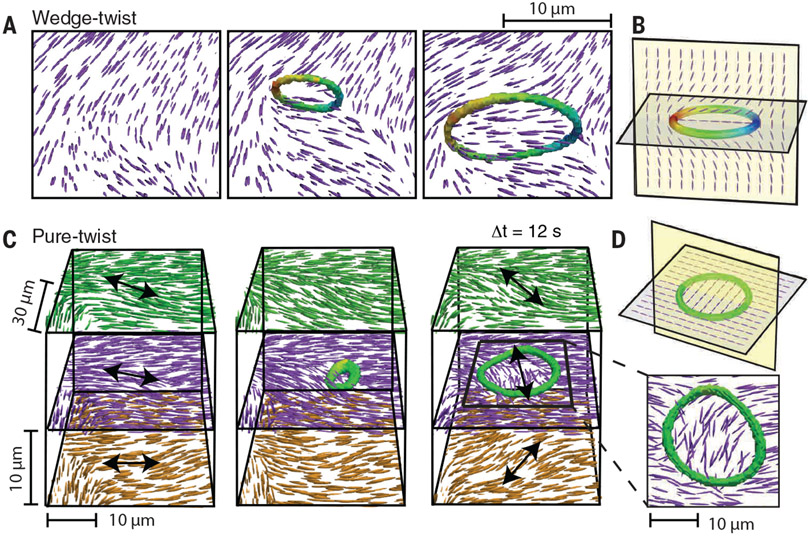
Topological structures are effective descriptors of the nonequilibrium dynamics of diverse many-body systems. For example, motile, point-like topological defects capture the salient features of two-dimensional active liquid crystals composed of energy-consuming anisotropic units. We dispersed force-generating microtubule bundles in a passive colloidal liquid crystal to form a three-dimensional active nematic. Light-sheet microscopy revealed the temporal evolution of the millimeter-scale structure of these active nematics with single-bundle resolution. The primary topological excitations are extended, charge-neutral disclination loops that undergo complex dynamics and recombination events. Our work suggests a framework for analyzing the nonequilibrium dynamics of bulk anisotropic systems as diverse as driven complex fluids, active metamaterials, biological tissues, and collections of robots or organisms.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures



Comment in
-
Rings rule three-dimensional active matter.Science. 2020 Mar 6;367(6482):1075-1076. doi: 10.1126/science.aba5319. Science. 2020. PMID: 32139530 No abstract available.
References
-
- Galtier S, Introduction to Modern Magnetohydrodynamics. (Cambridge University Press, ed. 1, 2016).
-
- Blanch-Mercader C et al. , Turbulent Dynamics of Epithelial Cell Cultures. Phys Rev Lett 120, 208101 (2018). - PubMed
-
- Bowick MJ, Chandar L, Schiff EA, Srivastava AM, The cosmological Kibble mechanism in the laboratory: string formation in liquid crystals. Science 263, 943–945 (1994). - PubMed
-
- Poulin P, Stark H, Lubensky T, Weitz D, Novel colloidal interactions in anisotropic fluids. Science 275, 1770–1773 (1997). - PubMed
-
- Bausch A et al. , Grain boundary scars and spherical crystallography. Science 299, 1716–1718 (2003). - PubMed
Publication types
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
