

Preferential HLA-B27 Allorecognition Displayed by Multiple Cross-Reactive Antiviral CD8+ T Cell Receptors
- PMID: 32140156
- PMCID: PMC7042382
- DOI: 10.3389/fimmu.2020.00248
Preferential HLA-B27 Allorecognition Displayed by Multiple Cross-Reactive Antiviral CD8+ T Cell Receptors
Abstract
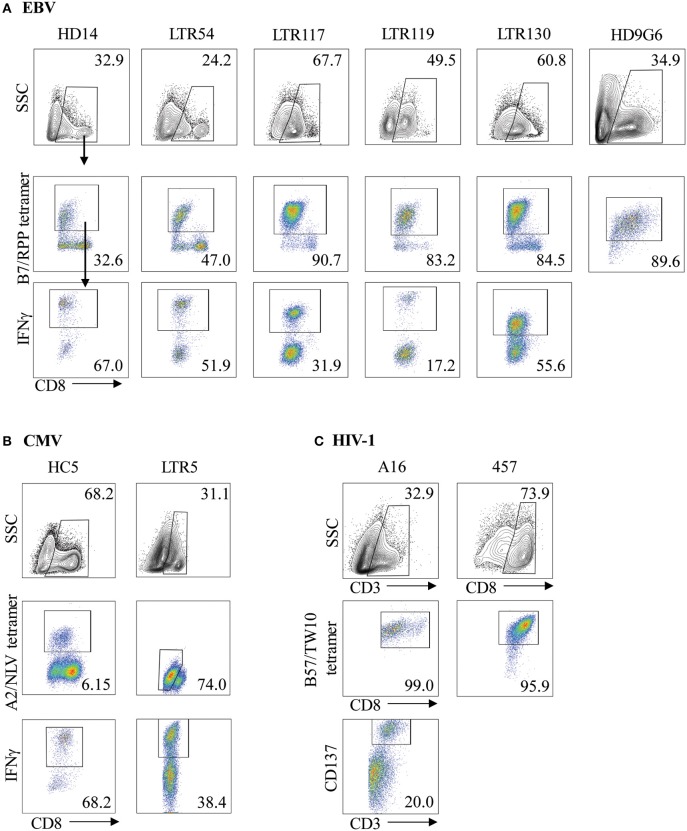
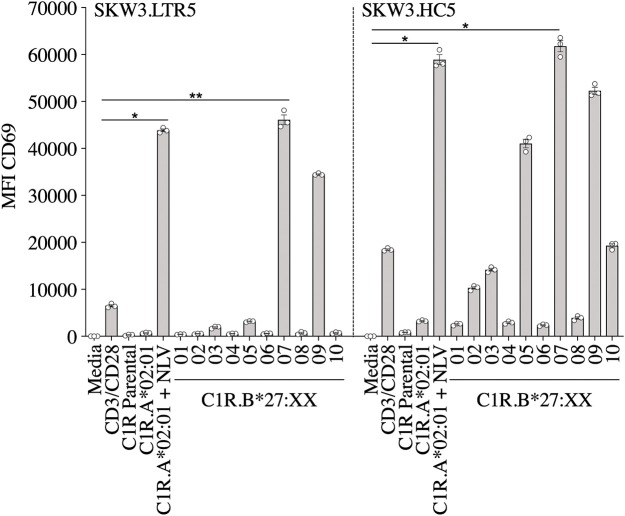
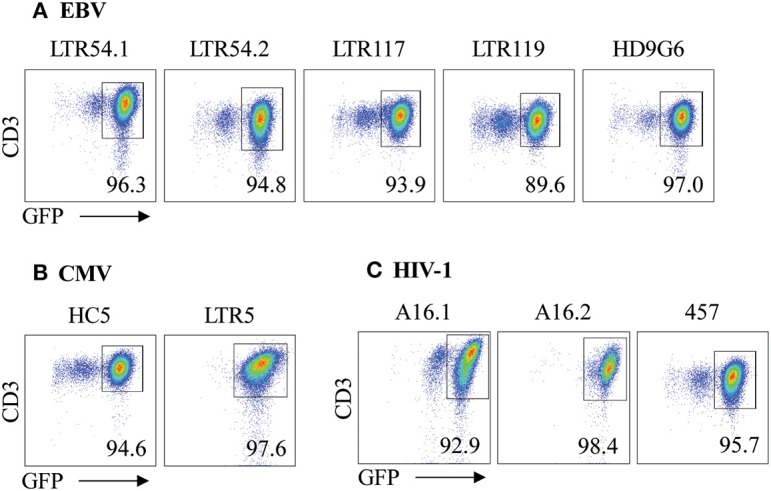
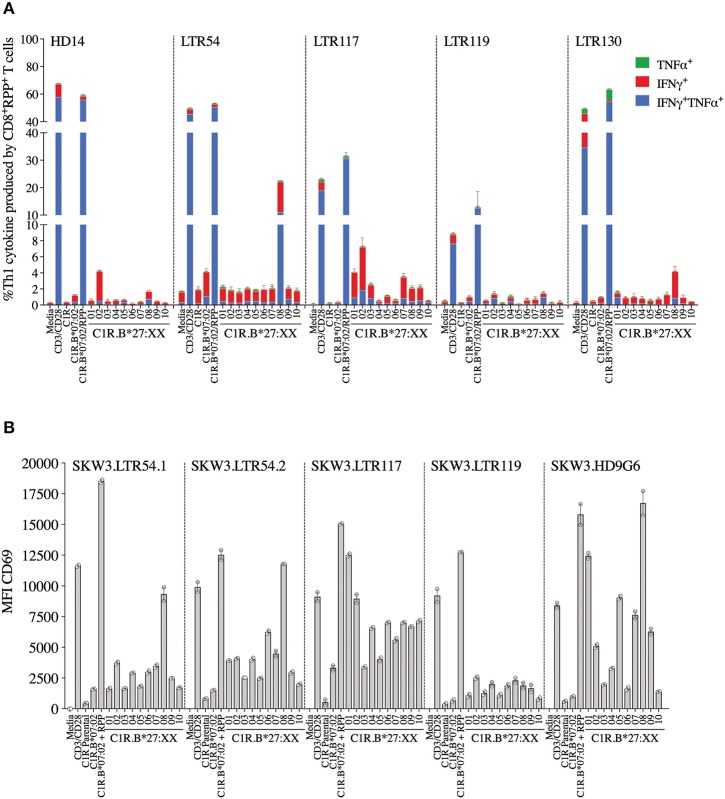
T cells provide essential immunosurveillance to combat and eliminate infection from pathogens, yet these cells can also induce unwanted immune responses via T cell receptor (TCR) cross-reactivity, also known as heterologous immunity. Indeed, pathogen-induced TCR cross-reactivity has shown to be a common, robust, and functionally potent mechanism that can trigger a spectrum of human immunopathologies associated with either transplant rejection, drug allergy, and autoimmunity. Here, we report that several virus-specific CD8+ T cells directed against peptides derived from chronic viruses (EBV, CMV, and HIV-1) presented by high frequency HLA-A and -B allomorphs differentially cross-react toward HLA-B27 allotypes in a highly focused and hierarchical manner. Given the commonality of cross-reactive T cells and their potential contribution to adverse outcomes in allogeneic transplants, our study demonstrates that multiple antiviral T cells recognizing the same HLA allomorph could pose an extra layer of complexity for organ matching.
Keywords: CMV; EBV; HIV-1; HLA; T cells; TCR; cross-reactivity.
Copyright © 2020 Rowntree, van den Heuvel, Sun, D'Orsogna, Nguyen, Claas, Rossjohn, Kotsimbos, Purcell and Mifsud.
Figures
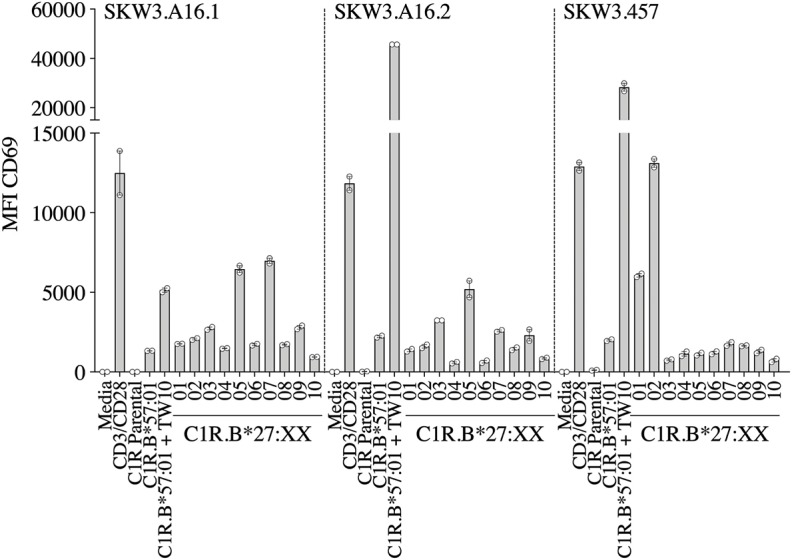
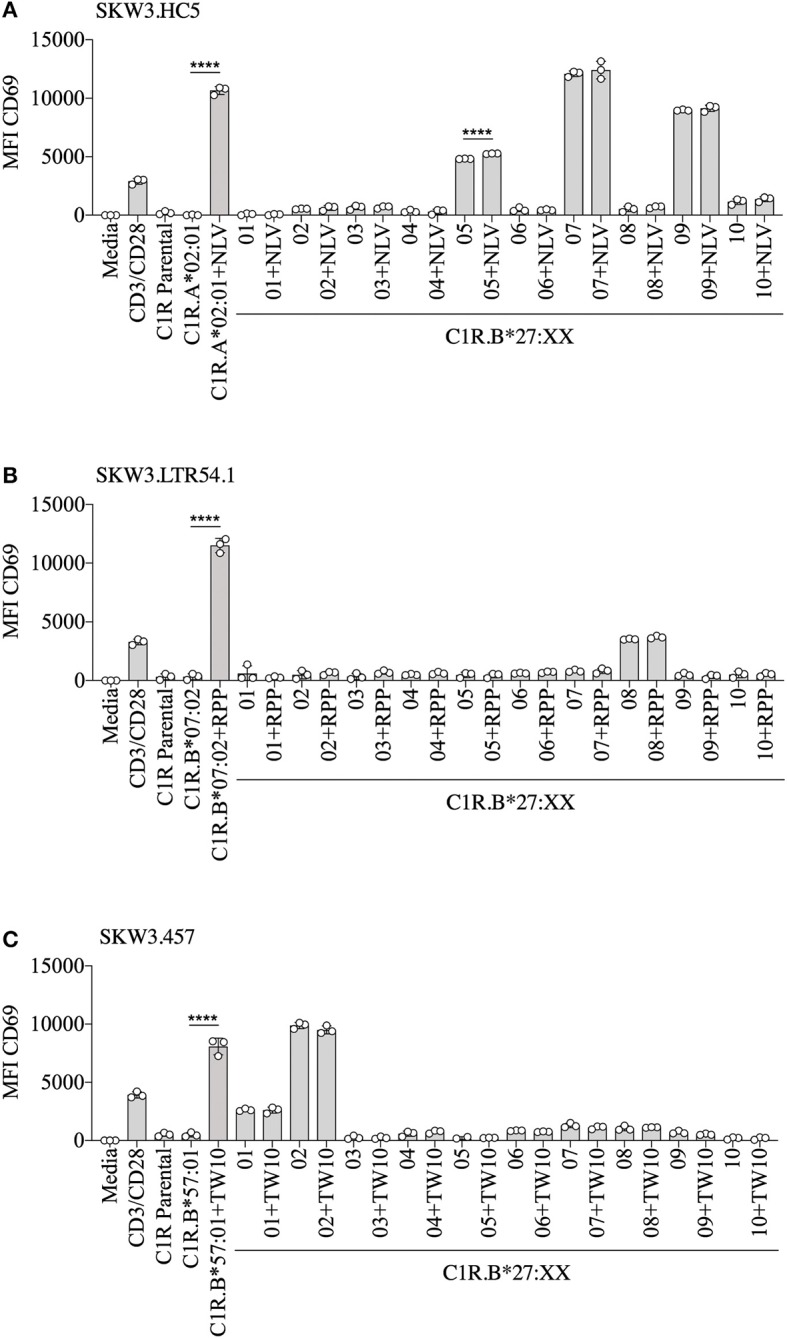
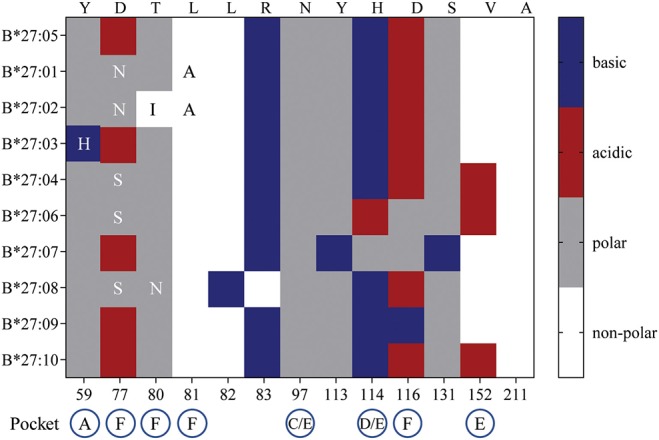
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials