

Genome-edited adult stem cells: Next-generation advanced therapy medicinal products
- PMID: 32141715
- PMCID: PMC7214650
- DOI: 10.1002/sctm.19-0338
Genome-edited adult stem cells: Next-generation advanced therapy medicinal products
Abstract
Over recent decades, gene therapy, which has enabled the treatment of several incurable diseases, has undergone a veritable revolution. Cell therapy has also seen major advances in the treatment of various diseases, particularly through the use of adult stem cells (ASCs). The combination of gene and cell therapy (GCT) has opened up new opportunities to improve advanced therapy medicinal products for the treatment of several diseases. Despite the considerable potential of GCT, the use of retroviral vectors has major limitations with regard to oncogene transactivation and the lack of physiological expression. Recently, gene therapists have focused on genome editing (GE) technologies as an alternative strategy. In this review, we discuss the potential benefits of using GE technologies to improve GCT approaches based on ASCs. We will begin with a brief summary of different GE platforms and techniques and will then focus on key therapeutic approaches that have been successfully used to treat diseases in animal models. Finally, we discuss whether ASC GE could become a real alternative to retroviral vectors in a GCT setting.
Keywords: CRISPR; adult stem cells; electroporation; gene delivery systems in vivo or in vitro; gene therapy; hematopoietic stem cells (HSCs); mesenchymal stem cells (MSCs); pluripotent hemopoietic stem cells.
© 2020 The Authors. Stem Cells Translational Medicine published by Wiley Periodicals, Inc. on behalf of AlphaMed Press.
Conflict of interest statement
The authors declared no potential conflicts of interest.
Figures
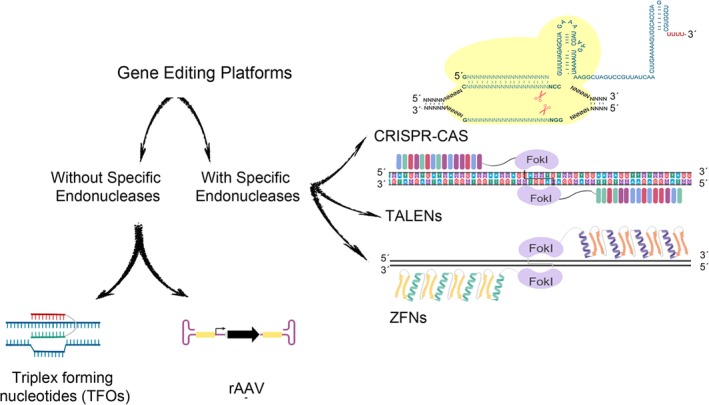
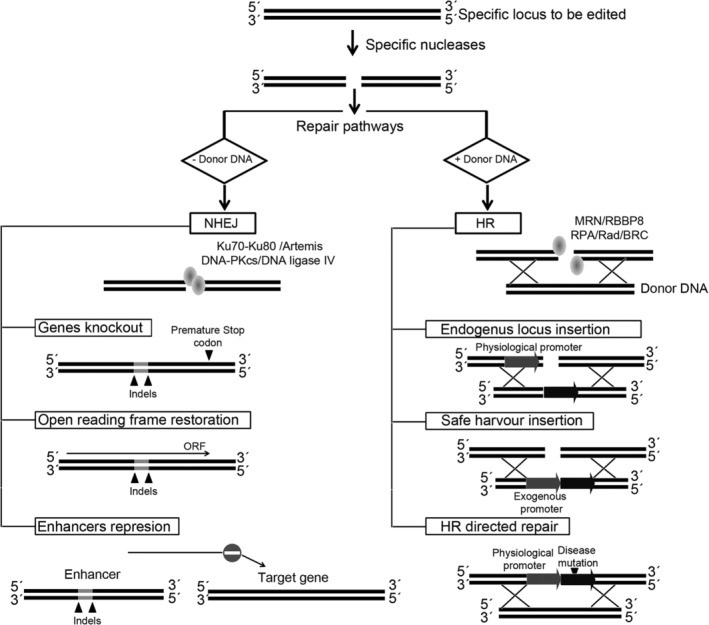
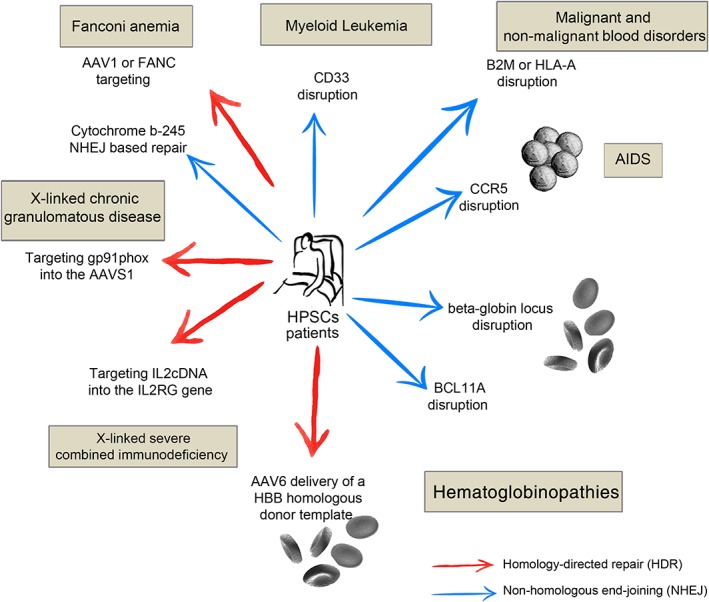
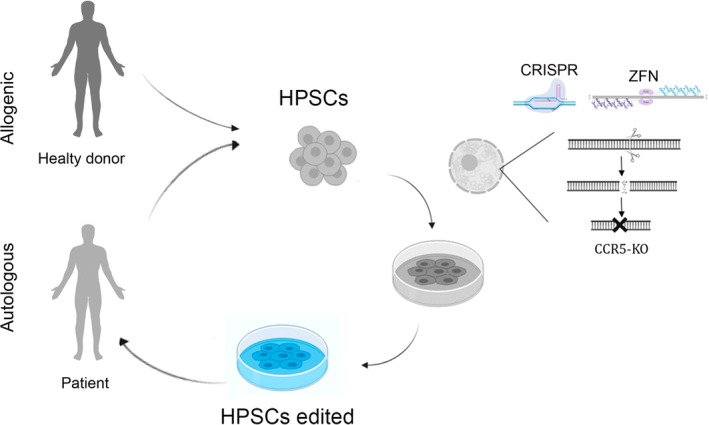
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
