

Mycobacterium tuberculosis Sulfolipid-1 Activates Nociceptive Neurons and Induces Cough
- PMID: 32142653
- PMCID: PMC7102531
- DOI: 10.1016/j.cell.2020.02.026
Mycobacterium tuberculosis Sulfolipid-1 Activates Nociceptive Neurons and Induces Cough
Abstract
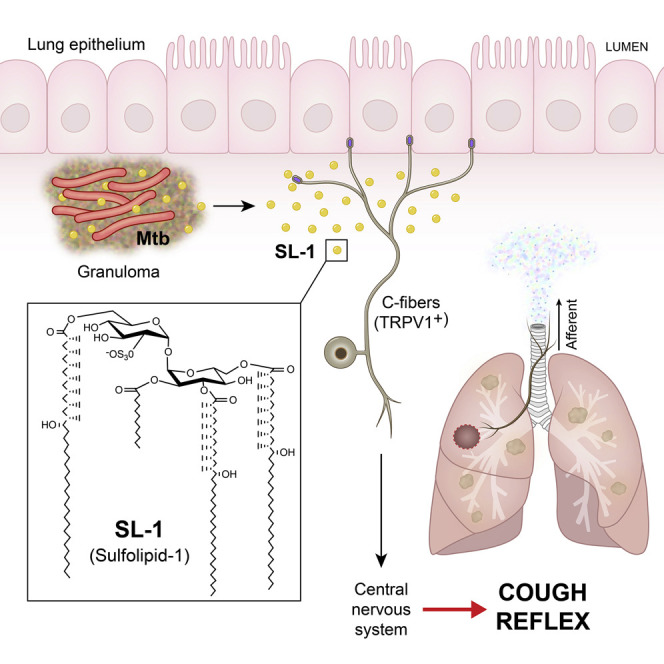
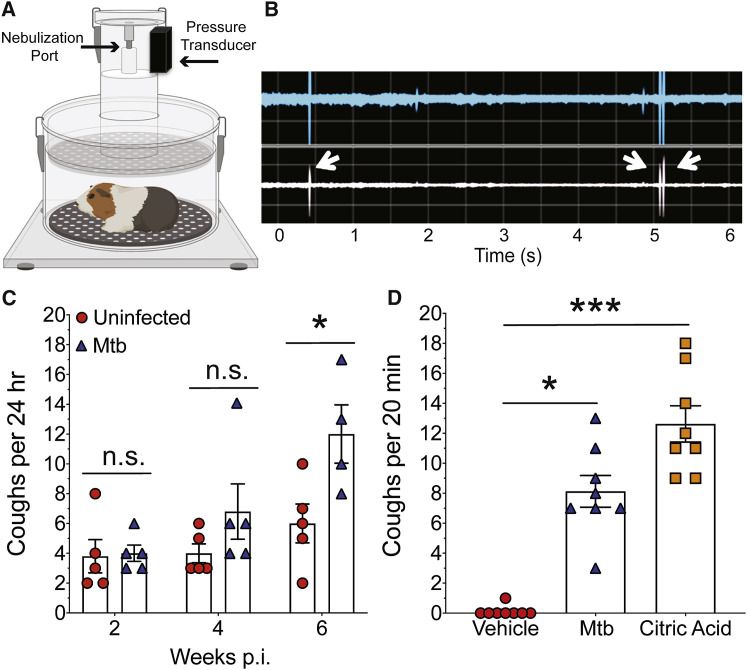
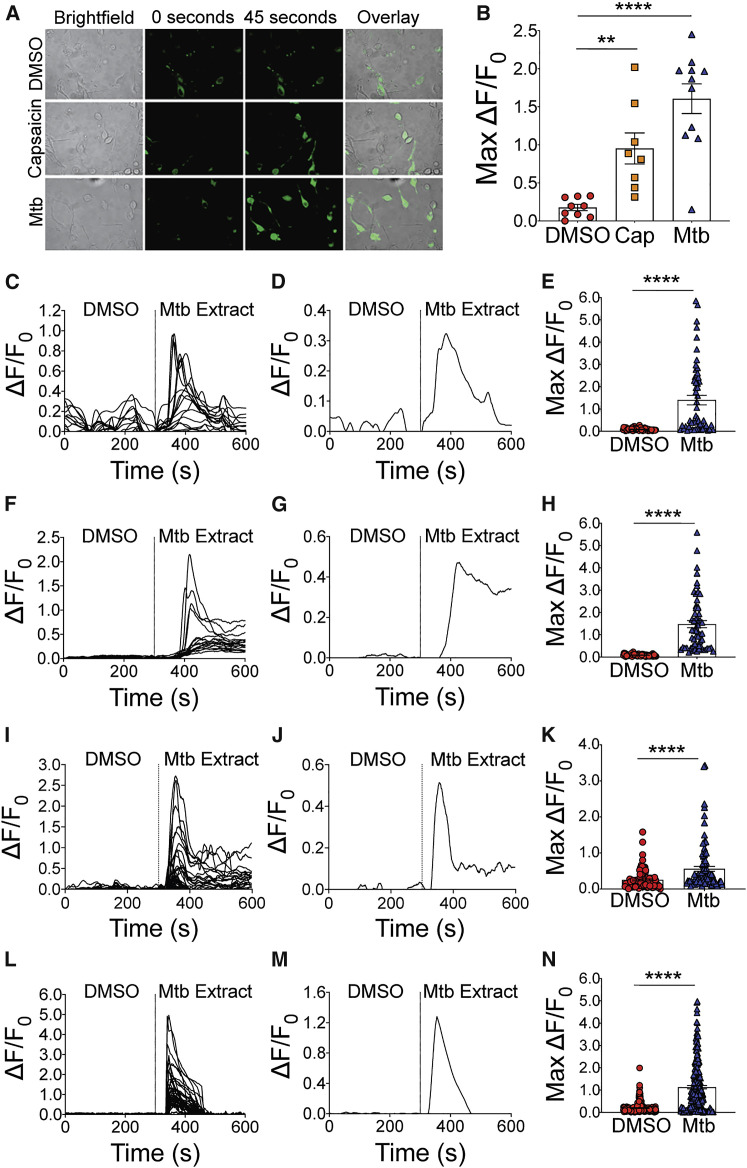
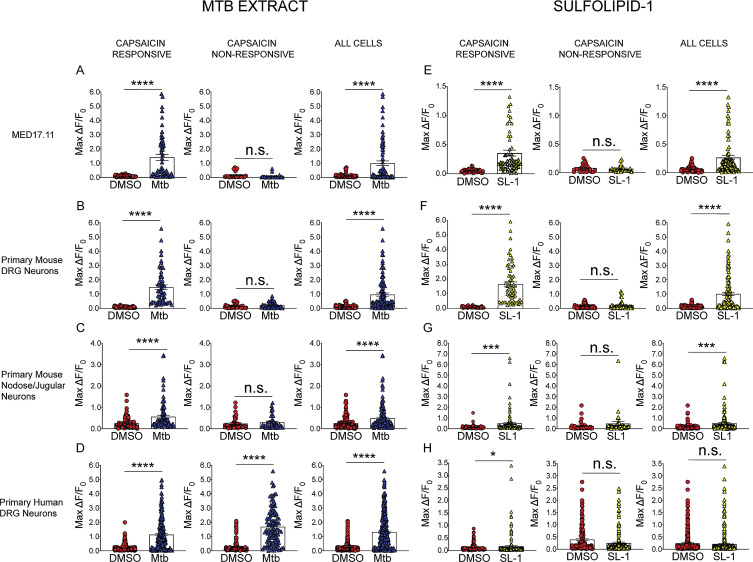
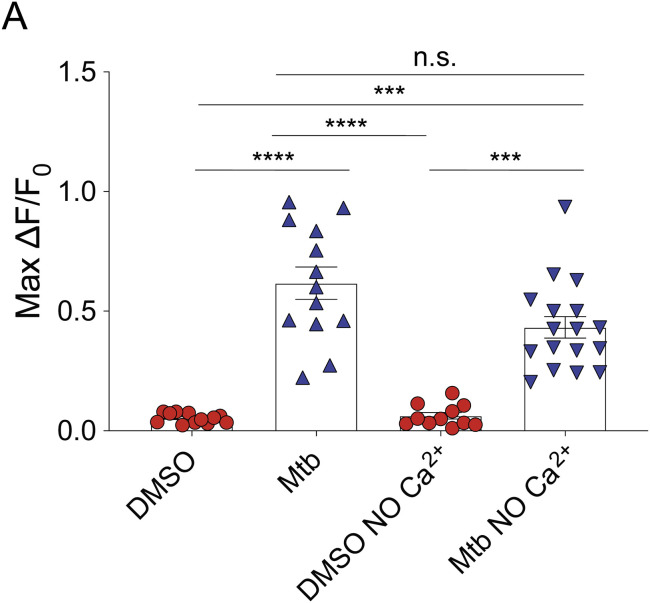
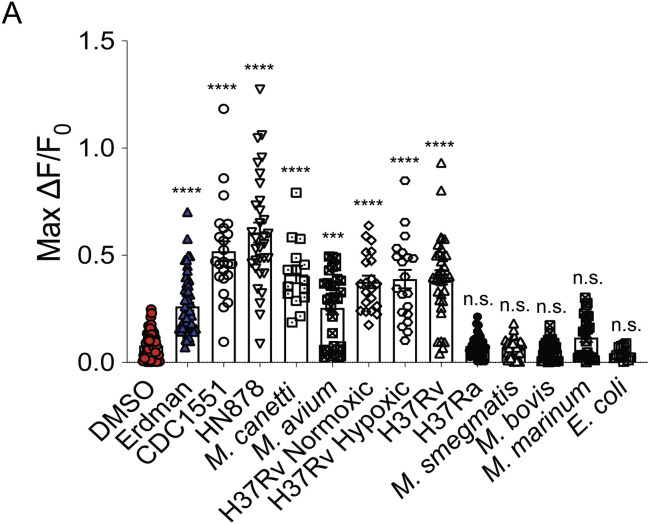
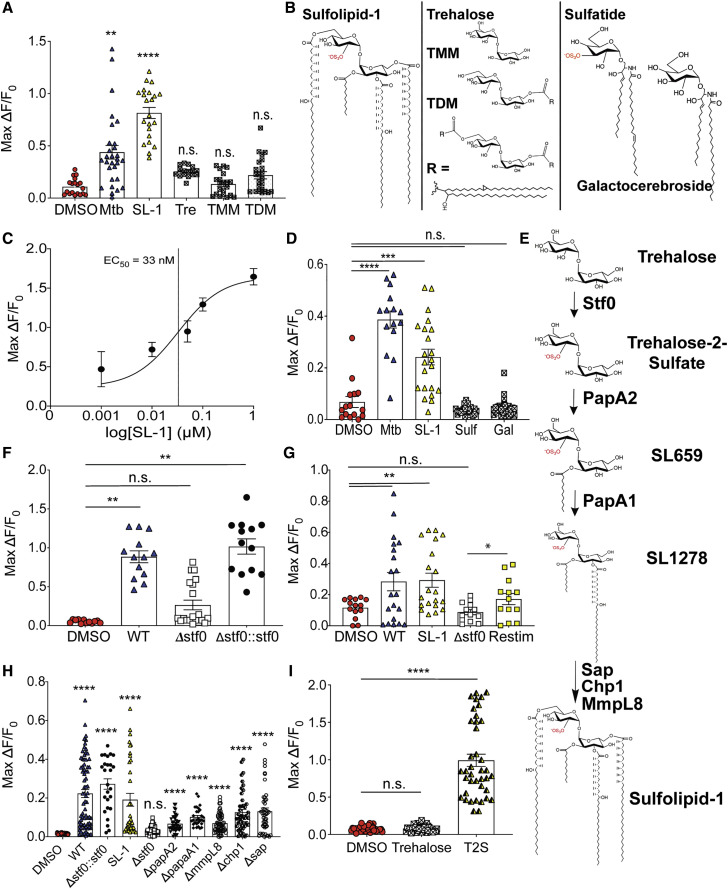
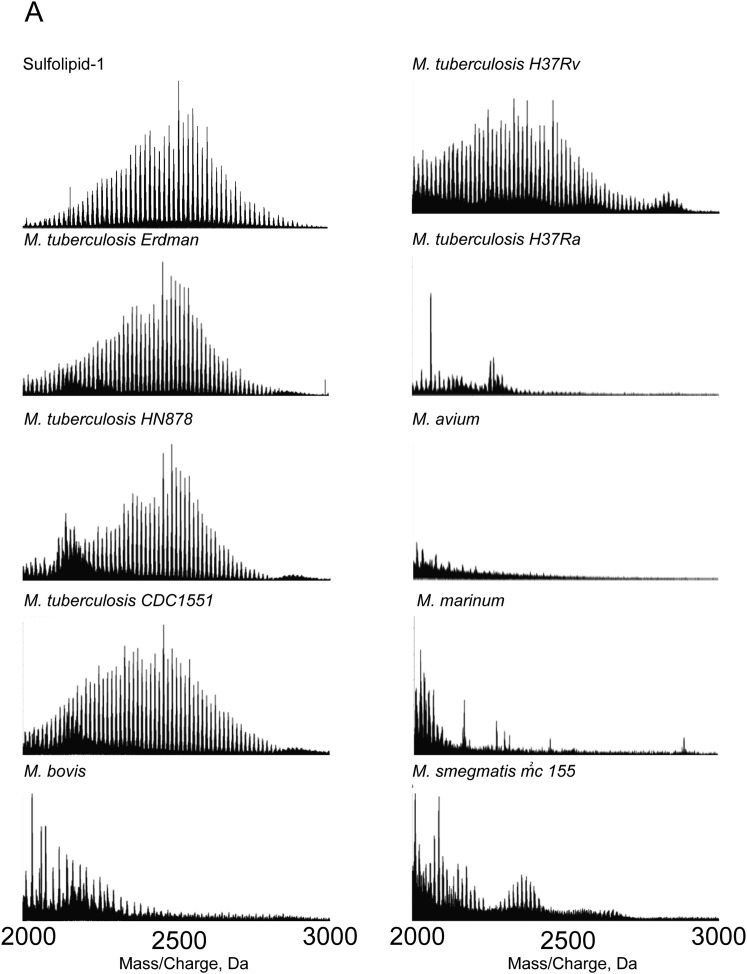
Pulmonary tuberculosis, a disease caused by Mycobacterium tuberculosis (Mtb), manifests with a persistent cough as both a primary symptom and mechanism of transmission. The cough reflex can be triggered by nociceptive neurons innervating the lungs, and some bacteria produce neuron-targeting molecules. However, how pulmonary Mtb infection causes cough remains undefined, and whether Mtb produces a neuron-activating, cough-inducing molecule is unknown. Here, we show that an Mtb organic extract activates nociceptive neurons in vitro and identify the Mtb glycolipid sulfolipid-1 (SL-1) as the nociceptive molecule. Mtb organic extracts from mutants lacking SL-1 synthesis cannot activate neurons in vitro or induce cough in a guinea pig model. Finally, Mtb-infected guinea pigs cough in a manner dependent on SL-1 synthesis. Thus, we demonstrate a heretofore unknown molecular mechanism for cough induction by a virulent human pathogen via its production of a complex lipid.
Keywords: cough; glycolipid; host-pathogen; mucosal immunology; mycobacteria; neuro-immune; nociceptor; sulfolipid; tuberculosis.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
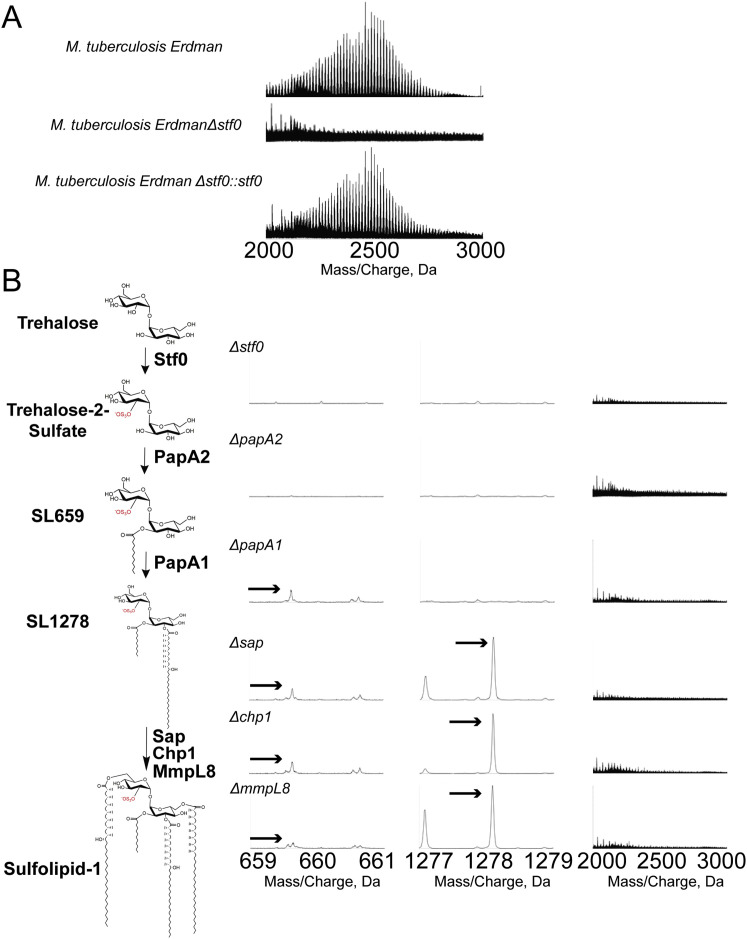
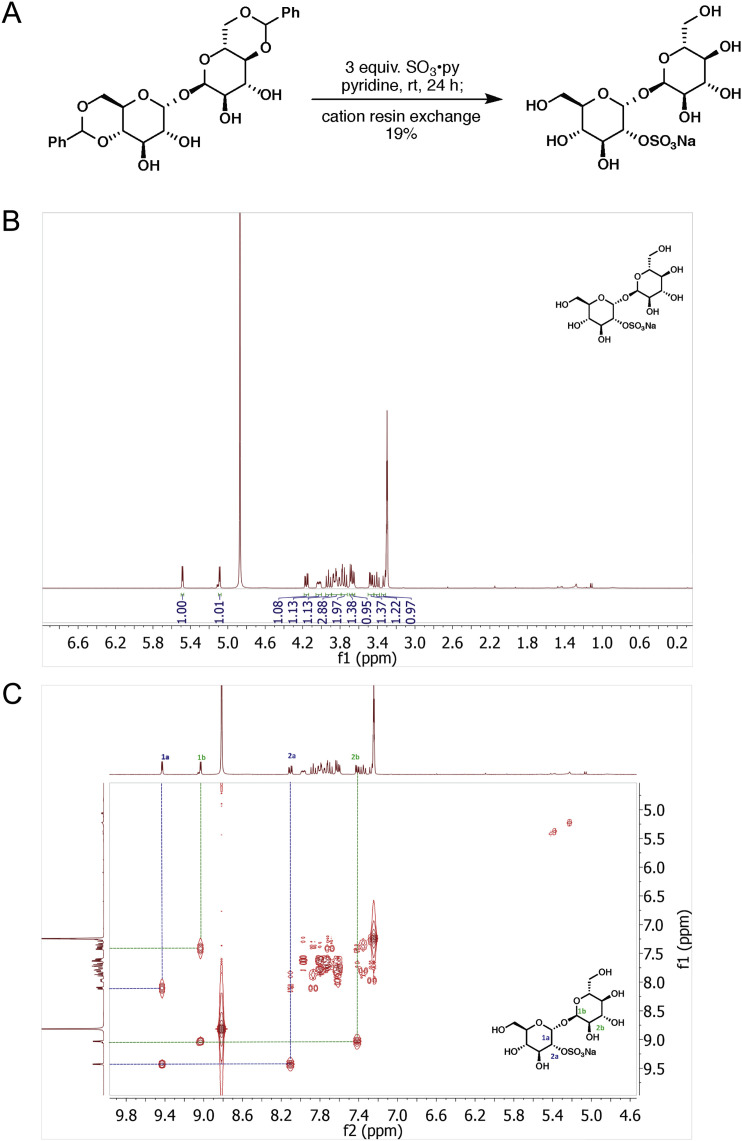
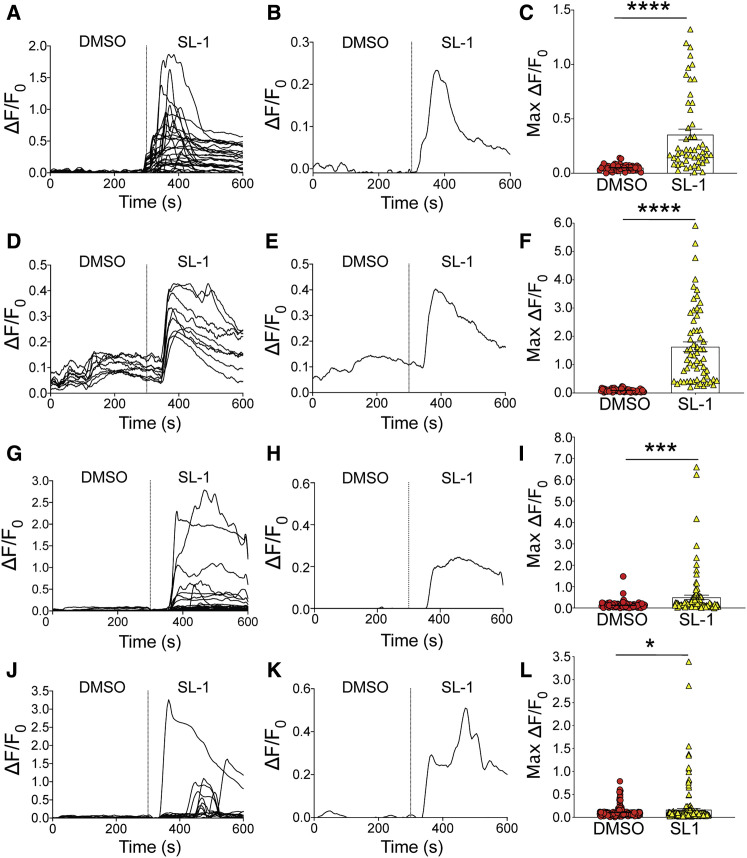
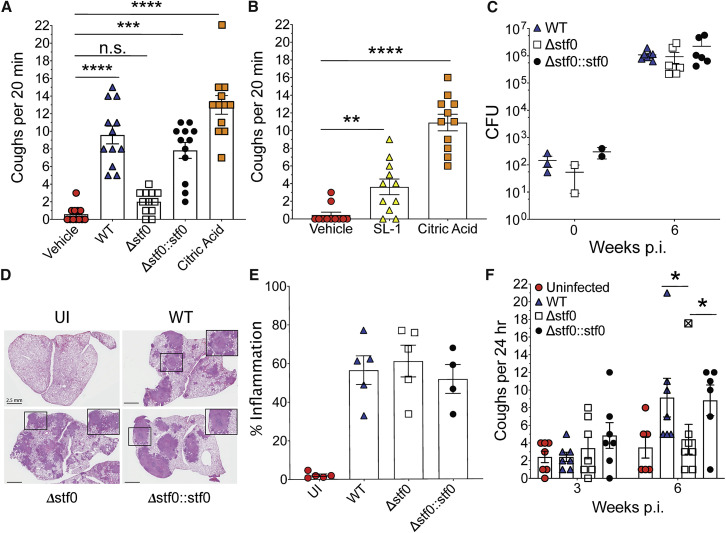
Comment in
-
SLeuthing Tuberculous Cough.Cell. 2020 Apr 16;181(2):230-232. doi: 10.1016/j.cell.2020.03.057. Cell. 2020. PMID: 32302566
References
-
- Abdullah H., Heaney L.G., Cosby S.L., McGarvey L.P. Rhinovirus upregulates transient receptor potential channels in a human neuronal cell line: implications for respiratory virus-induced cough reflex sensitivity. Thorax. 2014;69:46–54. - PubMed
-
- Abdulqawi R., Dockry R., Holt K., Layton G., McCarthy B.G., Ford A.P., Smith J.A. P2X3 receptor antagonist (AF-219) in refractory chronic cough: a randomised, double-blind, placebo-controlled phase 2 study. Lancet. 2015;385:1198–1205. - PubMed
-
- Basaraba R.J., Dailey D.D., McFarland C.T., Shanley C.A., Smith E.E., McMurray D.N., Orme I.M. Lymphadenitis as a major element of disease in the guinea pig model of tuberculosis. Tuberculosis (Edinb.) 2006;86:386–394. - PubMed
-
- Bonvini S.J., Belvisi M.G. Cough and airway disease: The role of ion channels. Pulm. Pharmacol. Ther. 2017;47:21–28. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
