

Biology of the human blood-nerve barrier in health and disease
- PMID: 32142802
- PMCID: PMC7145763
- DOI: 10.1016/j.expneurol.2020.113272
Biology of the human blood-nerve barrier in health and disease
Abstract
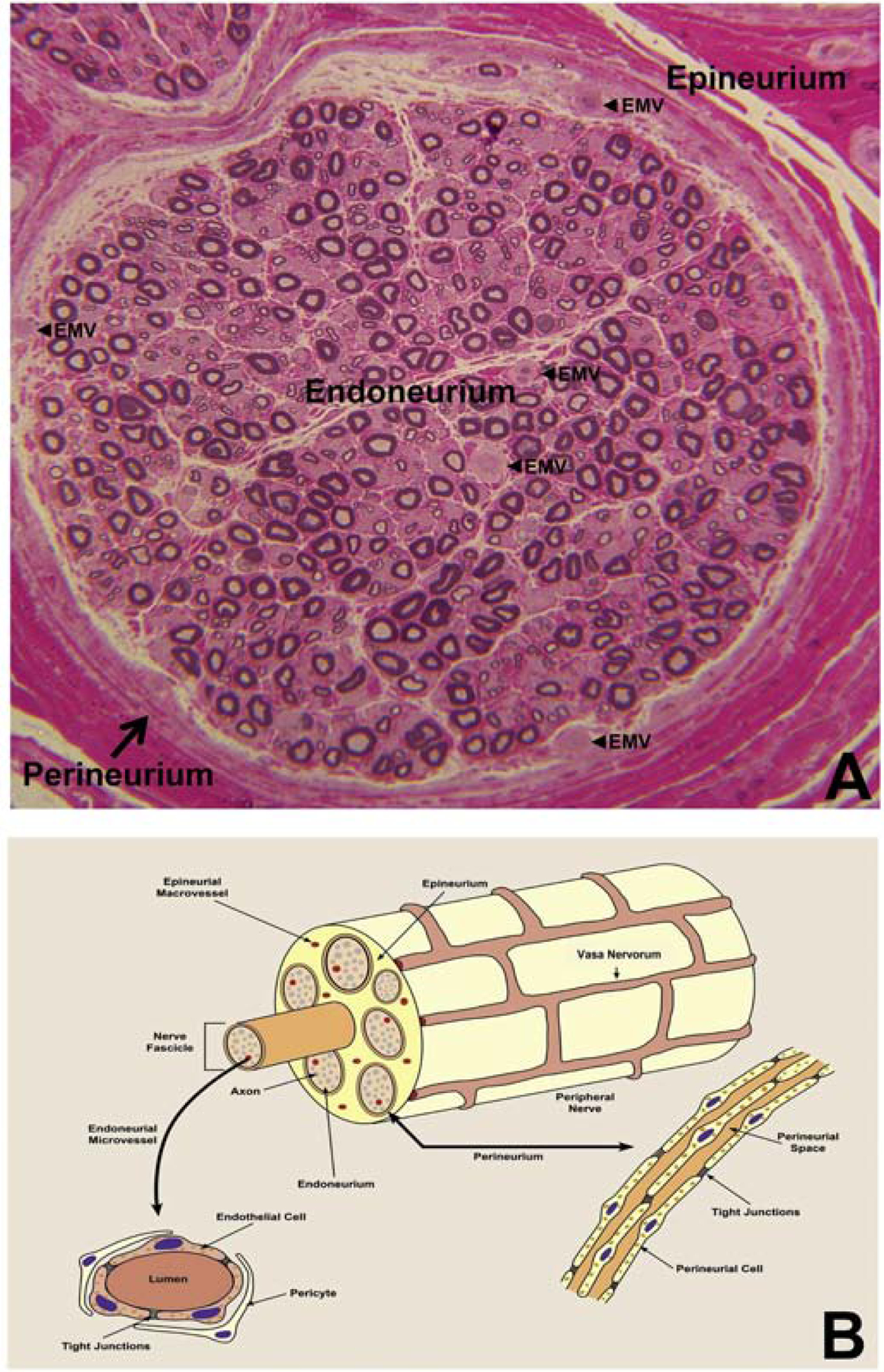
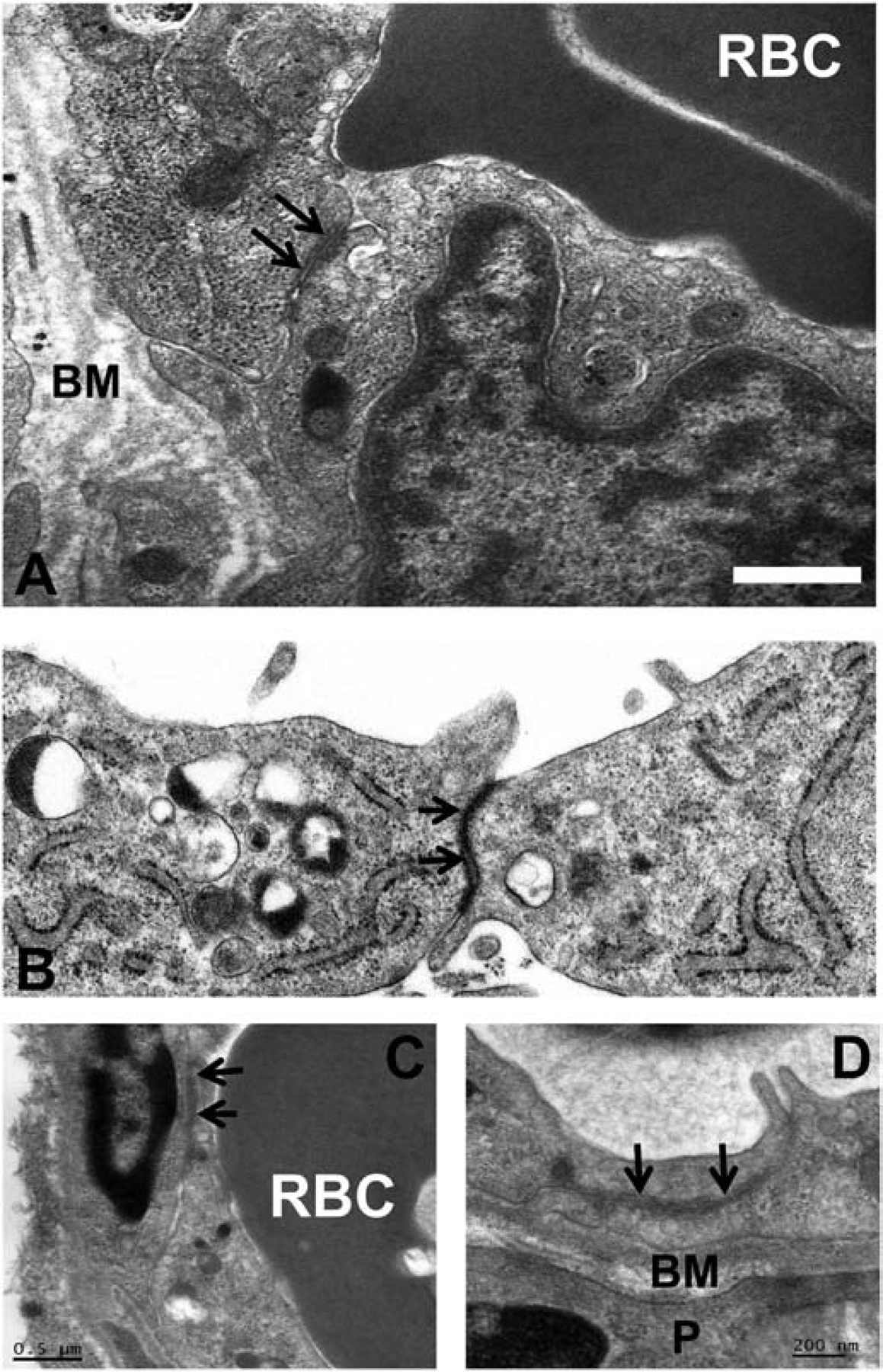
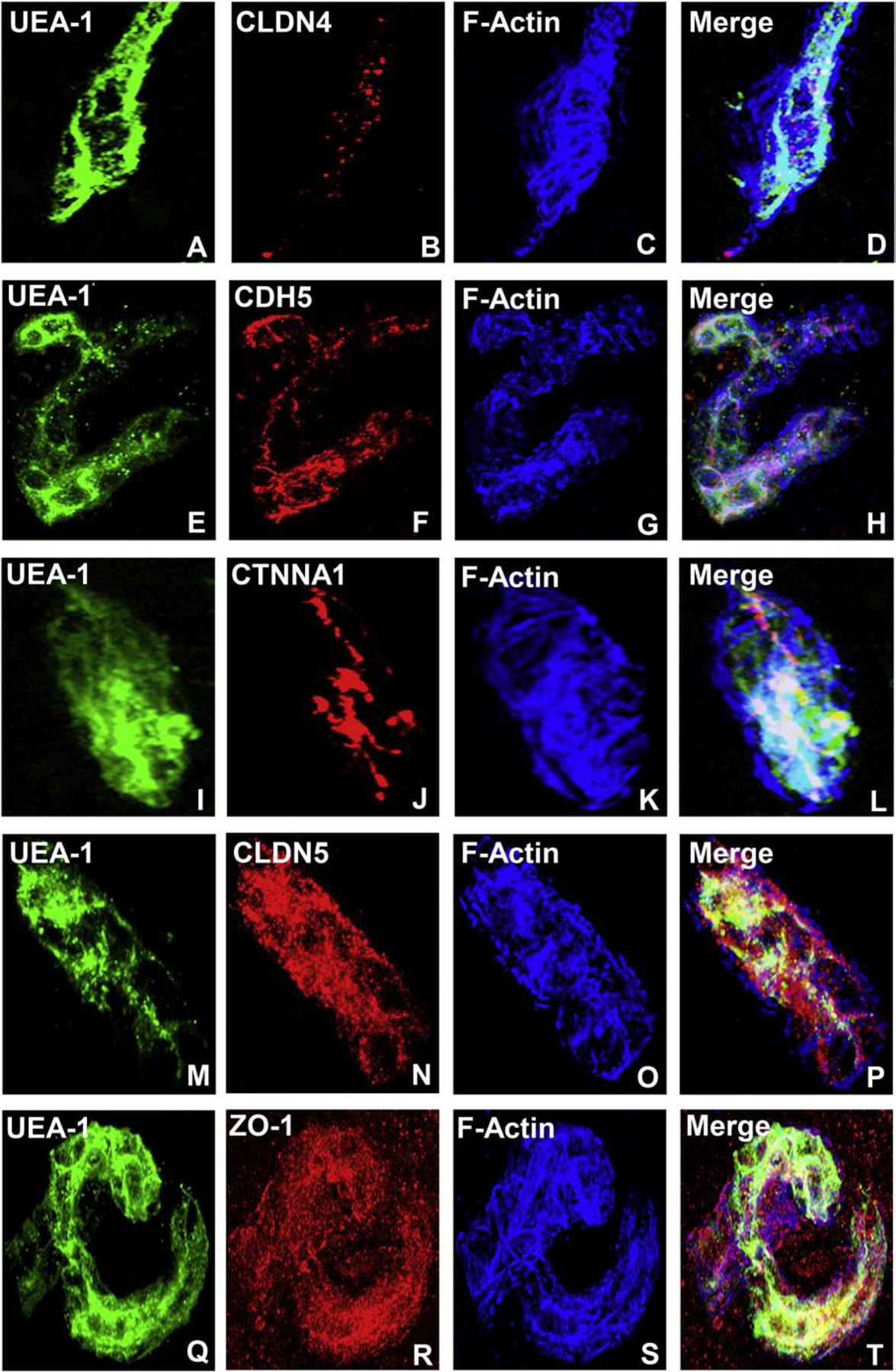
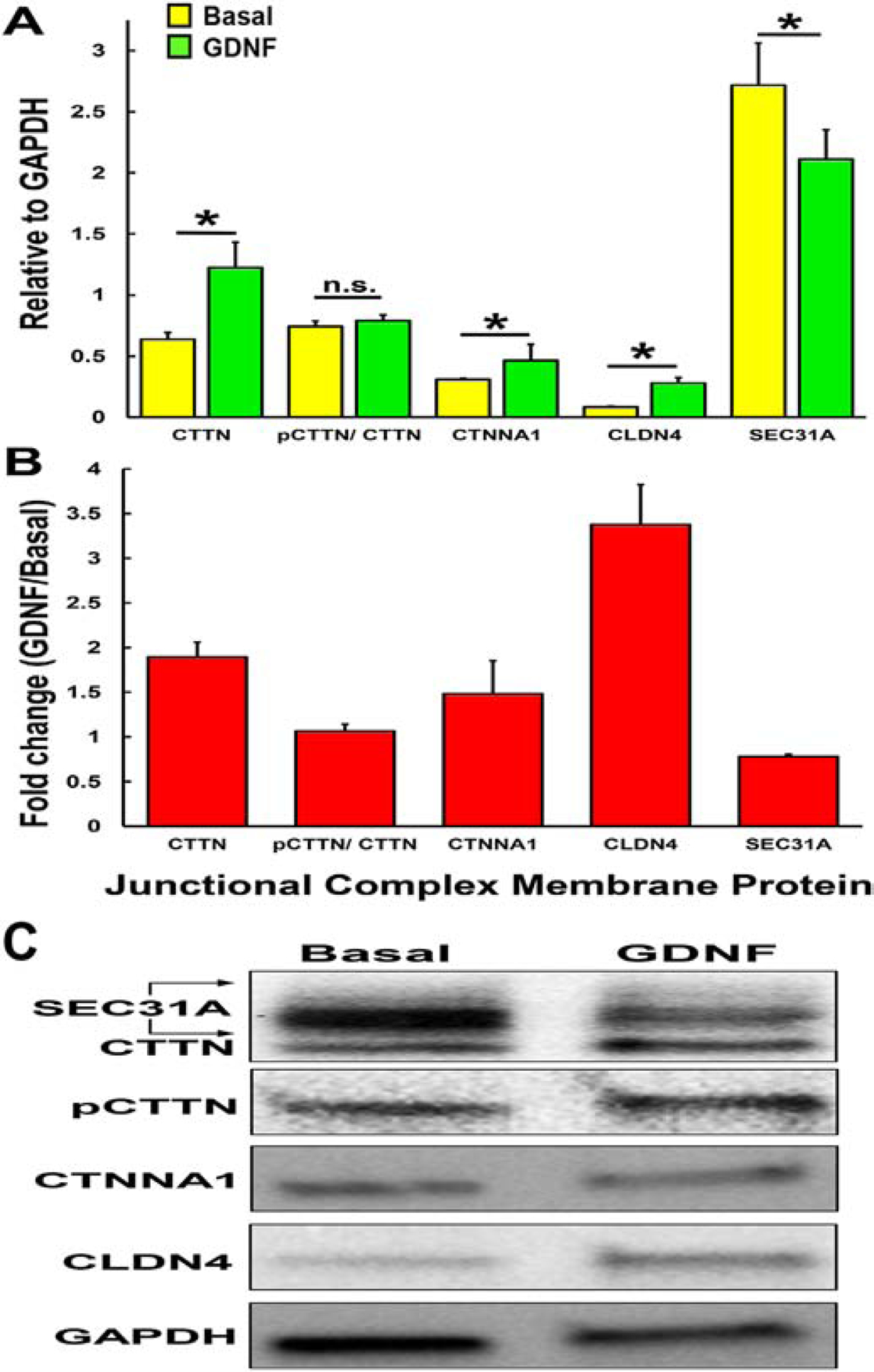
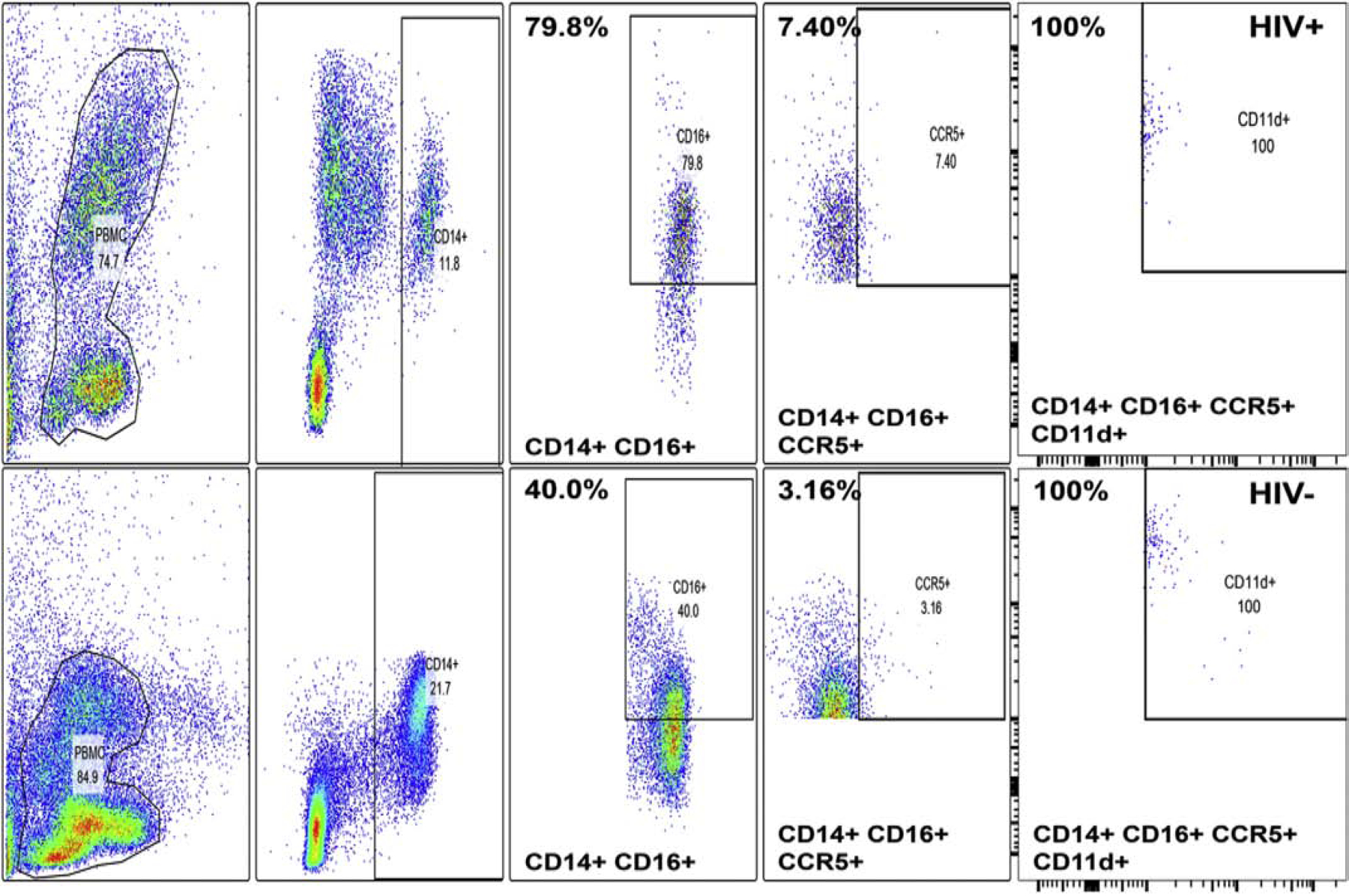
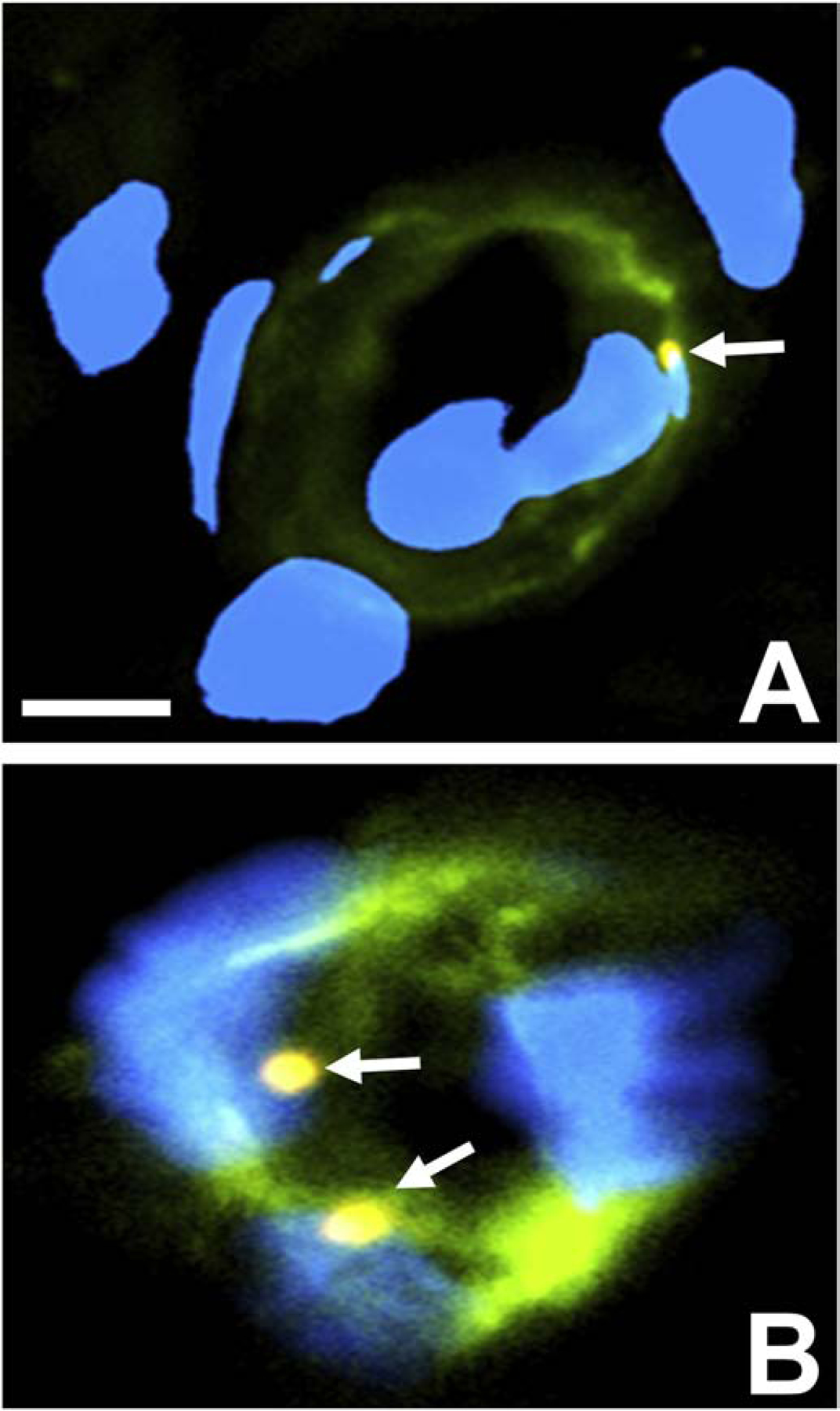
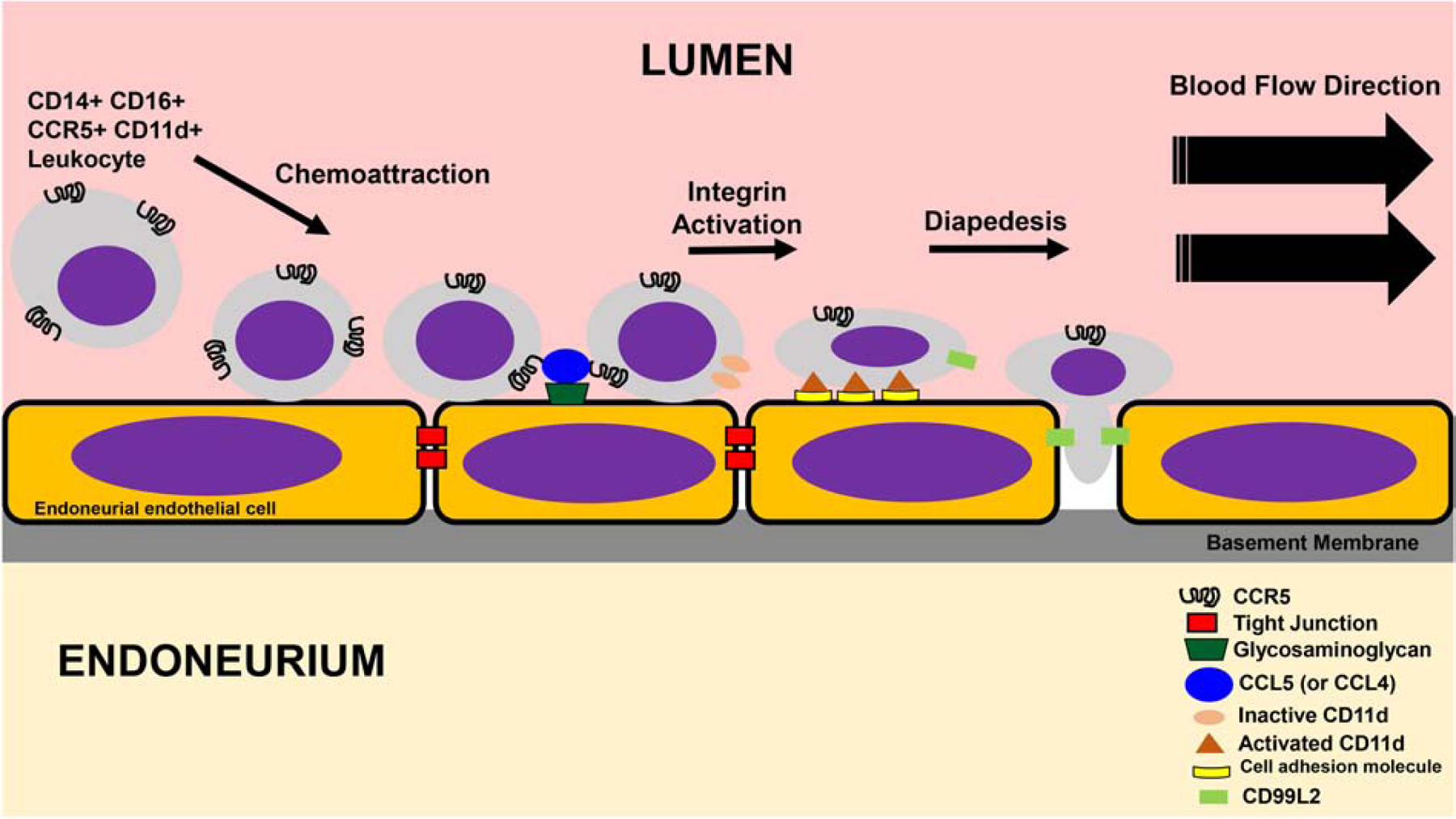
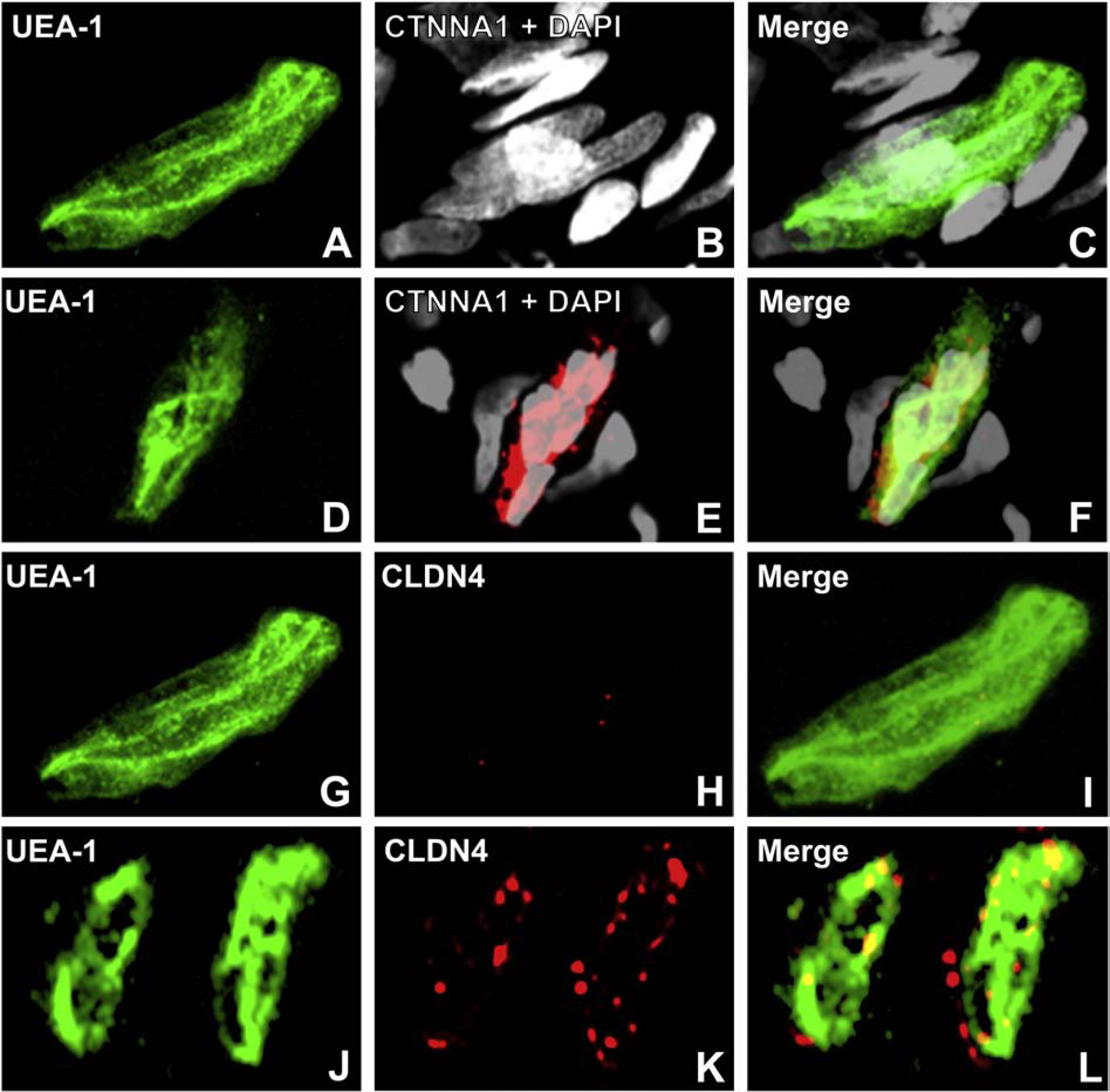
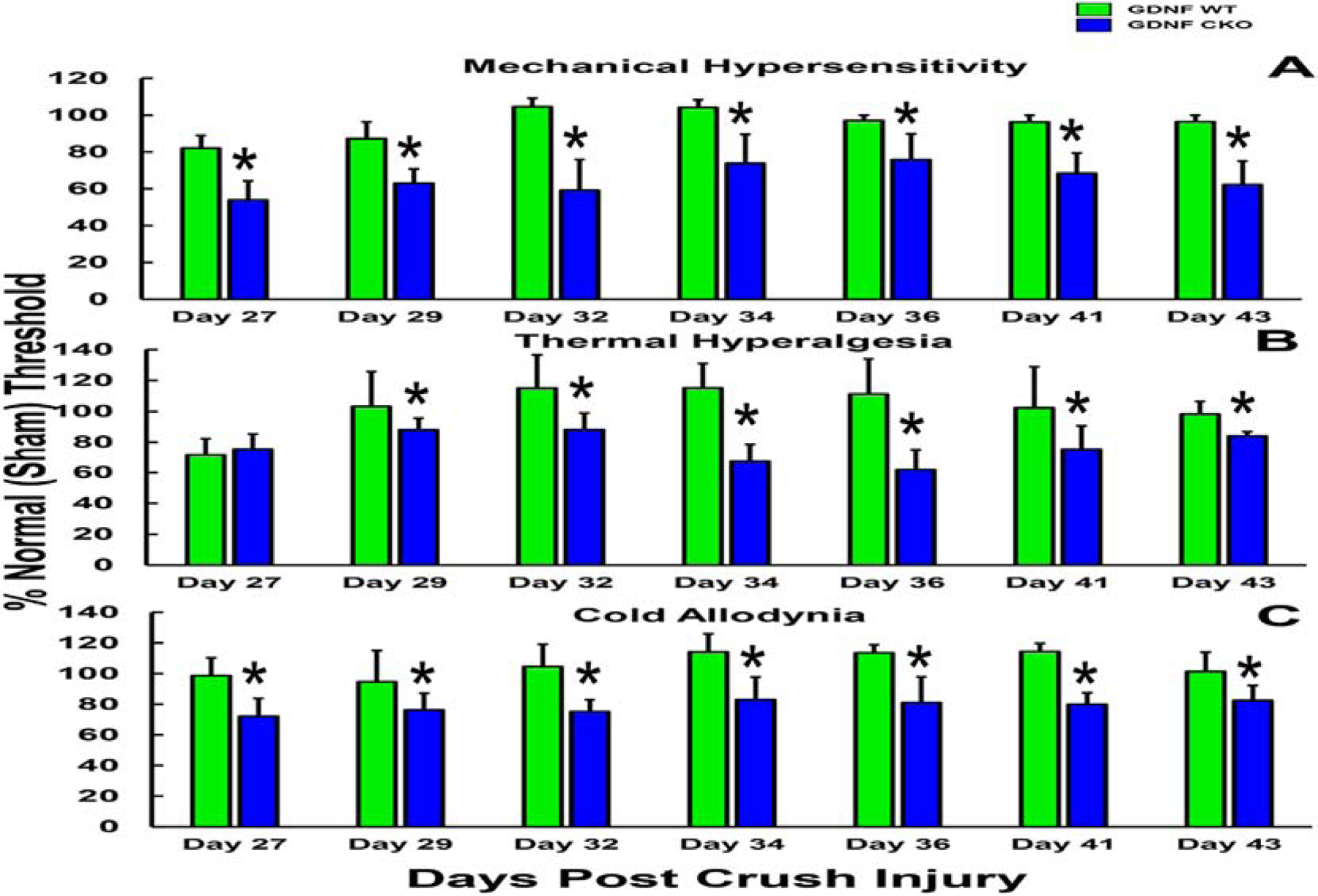
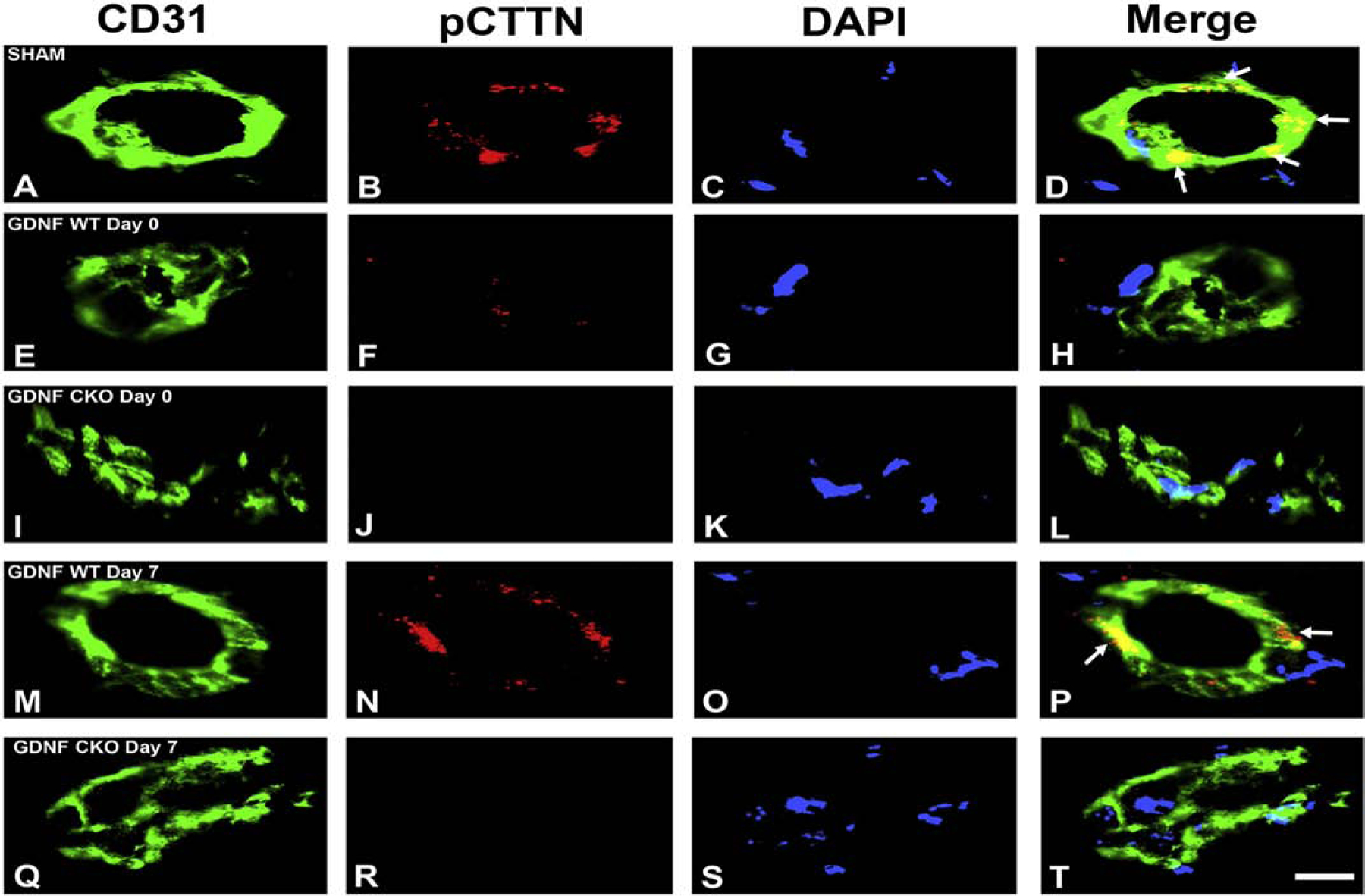
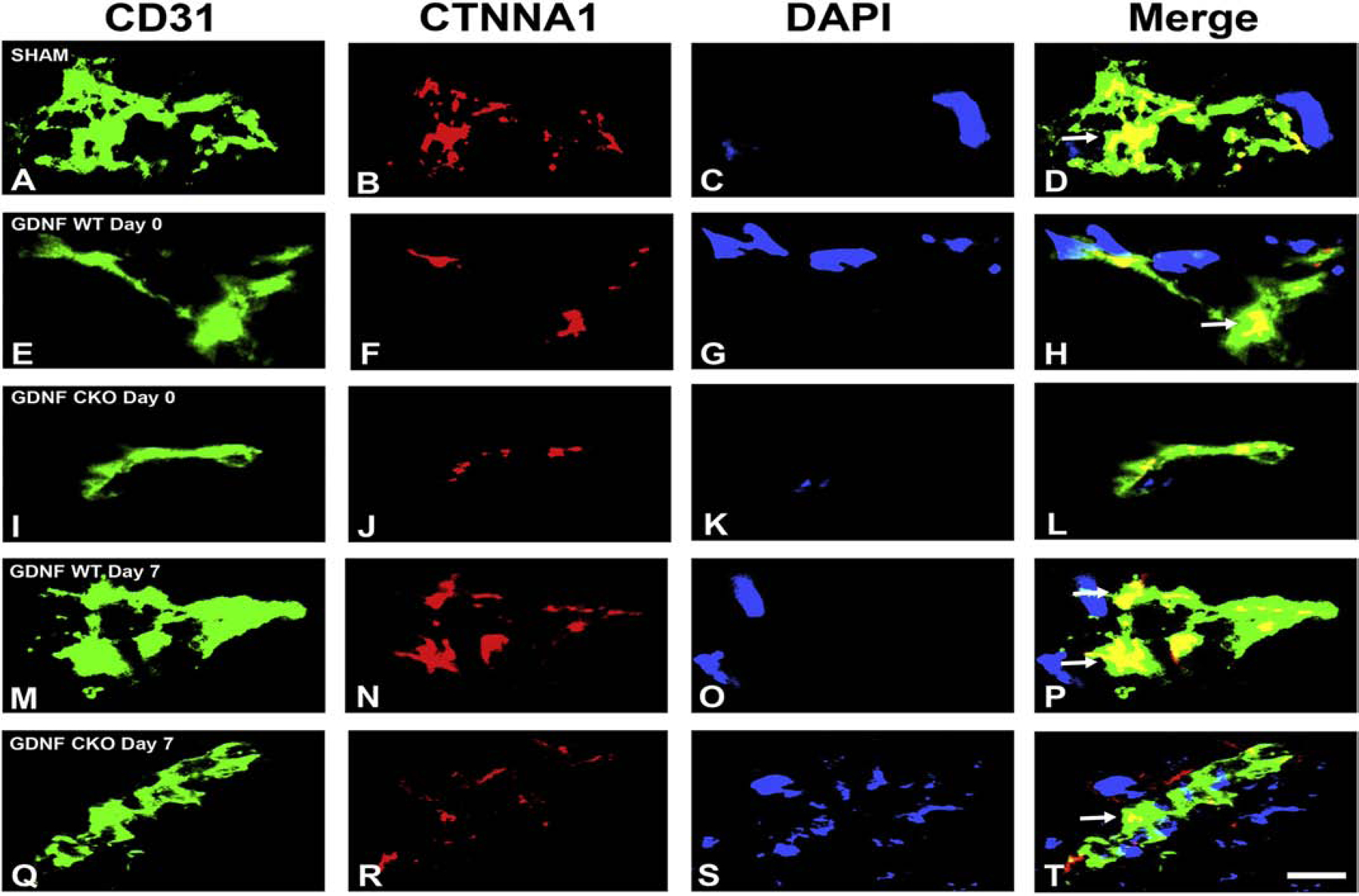
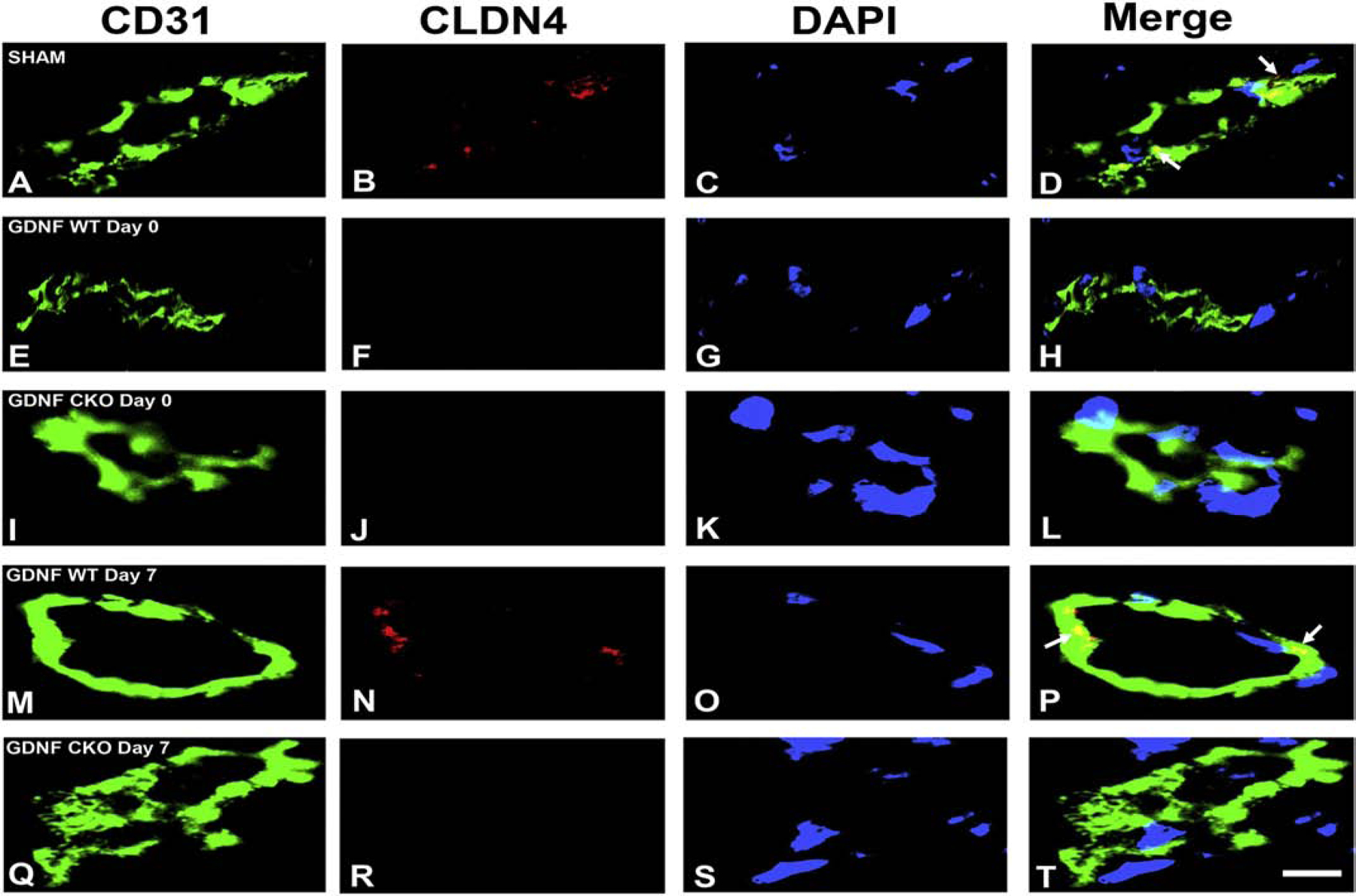
A highly regulated endoneurial microenvironment is required for normal axonal function in peripheral nerves and nerve roots, which structurally consist of an outer collagenous epineurium, inner perineurium consisting of multiple concentric layers of specialized epithelioid myofibroblasts that surround the innermost endoneurium, which consists of myelinated and unmyelinated axons embedded in a looser mesh of collagen fibers. Endoneurial homeostasis is achieved by tight junction-forming endoneurial microvessels that control ion, solute, water, nutrient, macromolecule and leukocyte influx and efflux between the bloodstream and endoneurium, and the innermost layers of the perineurium that control interstitial fluid component flux between the freely permeable epineurium and endoneurium. Strictly speaking, endoneurial microvascular endothelium should be considered the blood-nerve barrier (BNB) due to direct communication with circulating blood. The mammalian BNB is considered the second most restrictive vascular system after the blood-brain barrier (BBB) based on classic in situ permeability studies. Structural alterations in endoneurial microvessels or interactions with hematogenous leukocytes have been described in several human peripheral neuropathies; however major advances in BNB biology in health and disease have been limited over the past 50 years. Guided by transcriptome and proteome studies of normal and pathologic human peripheral nerves, purified primary and immortalized human endoneurial endothelial cells that form the BNB and leukocytes from patients with well-characterized peripheral neuropathies, validated by in situ or ex vivo protein expression studies, data are emerging on the molecular and functional characteristics of the human BNB in health and in specific peripheral neuropathies, as well as chronic neuropathic pain. These early advancements have the potential to not only increase our understanding of how the BNB works and adapts or fails to adapt to varying insult, but provide insights relevant to pathogenic leukocyte trafficking, with translational potential and specific therapeutic application for chronic peripheral neuropathies and neuropathic pain.
Keywords: Blood-nerve barrier; Chronic neuropathic pain; Endoneurium; Immune system; Inflammation; Junctional complex; Leukocyte trafficking; Peripheral neuropathy.
Copyright © 2020 Elsevier Inc. All rights reserved.
Figures
References
-
- Abe M, Sano Y, Maeda T, Shimizu F, Kashiwamura Y, Haruki H, Saito K, Tasaki A, Kawai M, Terasaki T, Kanda T, 2012. Establishment and characterization of human peripheral nerve microvascular endothelial cell lines: a new in vitro blood-nerve barrier (BNB) model. Cell Struct Funct 37, 89–100. - PubMed
-
- Allen SJ, Watson JJ, Shoemark DK, Barua NU, Patel NK, 2013. GDNF, NGF and BDNF as therapeutic options for neurodegeneration. Pharmacol Ther 138, 155–175. - PubMed
-
- Allt G, Lawrenson JG, 2000. The blood-nerve barrier: enzymes, transporters and receptors--a comparison with the blood-brain barrier. Brain Res Bull 52, 1–12. - PubMed
-
- Aziz MH, Cui K, Das M, Brown KE, Ardell CL, Febbraio M, Pluskota E, Han J, Wu H, Ballantyne CM, Smith JD, Cathcart MK, Yakubenko VP, 2017. The Upregulation of Integrin alphaDbeta2 (CD11d/CD18) on Inflammatory Macrophages Promotes Macrophage Retention in Vascular Lesions and Development of Atherosclerosis. J Immunol 198, 4855–4867. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
