

Evolving Always-Critical Networks
- PMID: 32143532
- PMCID: PMC7151631
- DOI: 10.3390/life10030022
Evolving Always-Critical Networks
Abstract
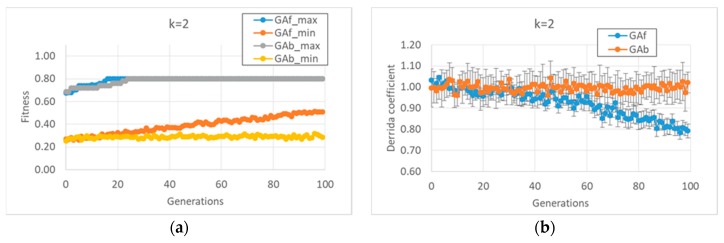
Living beings share several common features at the molecular level, but there are very few large-scale "operating principles" which hold for all (or almost all) organisms. However, biology is subject to a deluge of data, and as such, general concepts such as this would be extremely valuable. One interesting candidate is the "criticality" principle, which claims that biological evolution favors those dynamical regimes that are intermediaries between ordered and disordered states (i.e., "at the edge of chaos"). The reasons why this should be the case and experimental evidence are briefly discussed, observing that gene regulatory networks are indeed often found on, or close to, the critical boundaries. Therefore, assuming that criticality provides an edge, it is important to ascertain whether systems that are critical can further evolve while remaining critical. In order to explore the possibility of achieving such "always-critical" evolution, we resort to simulated evolution, by suitably modifying a genetic algorithm in such a way that the newly-generated individuals are constrained to be critical. It is then shown that these modified genetic algorithms can actually develop critical gene regulatory networks with two interesting (and quite different) features of biological significance, involving, in one case, the average gene activation values and, in the other case, the response to perturbations. These two cases suggest that it is often possible to evolve networks with interesting properties without losing the advantages of criticality. The evolved networks also show some interesting features which are discussed.
Keywords: Boolean models; criticality; edge of chaos; evolving systems; gene regulatory networks; genetic algorithms; random Boolean networks.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures




References
-
- Darwin C.R. On the Origin of Species. John Murray; London, UK: 1859.
-
- Kauffman S.A. The Origins of Order: Self-Organization and Selection in Evolution. Oxford University Press; New York, NY, USA: 1993.
-
- Kauffman S.A. At Home in the Universe. Oxford University Press; New York, NY, USA: 1995.
-
- Crutchfield J.P., Young K. Computation at the Onset of Chaos. In: Zurek W., editor. Entropy, Complexity, and the Physics of Information. Addison-Wesley; Reading, MA, USA: 1990. pp. 23–269. SFI Studies in the Sciences of Complexity, VIII.
-
- Langton C.G. Computation at the edge of chaos: Phase transitions and emergent computation. Phys. D Nonlinear Phenom. 1990;42:12–37. doi: 10.1016/0167-2789(90)90064-V. - DOI
LinkOut - more resources
Full Text Sources
