

Inhibition of alpha-synuclein seeded fibril formation and toxicity by herbal medicinal extracts
- PMID: 32143619
- PMCID: PMC7076823
- DOI: 10.1186/s12906-020-2849-1
Inhibition of alpha-synuclein seeded fibril formation and toxicity by herbal medicinal extracts
Retraction in
-
Retraction Note: Inhibition of alpha-synuclein seeded fibril formation and toxicity by herbal medicinal extracts.BMC Complement Med Ther. 2025 Nov 8;25(1):421. doi: 10.1186/s12906-025-05176-3. BMC Complement Med Ther. 2025. PMID: 41206449 Free PMC article. No abstract available.
Abstract
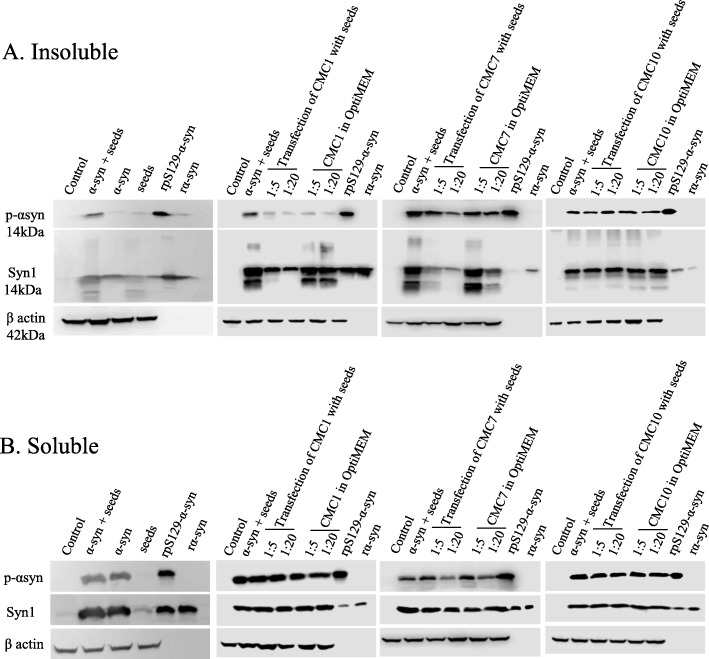
Background: Recent studies indicated that seeded fibril formation and toxicity of α-synuclein (α-syn) play a main role in the pathogenesis of certain diseases including Parkinson's disease (PD), multiple system atrophy, and dementia with Lewy bodies. Therefore, examination of compounds that abolish the process of seeding is considered a key step towards therapy of several synucleinopathies.
Methods: Using biophysical, biochemical and cell-culture-based assays, assessment of eleven compounds, extracted from Chinese medicinal herbs, was performed in this study for their effect on α-syn fibril formation and toxicity caused by the seeding process.
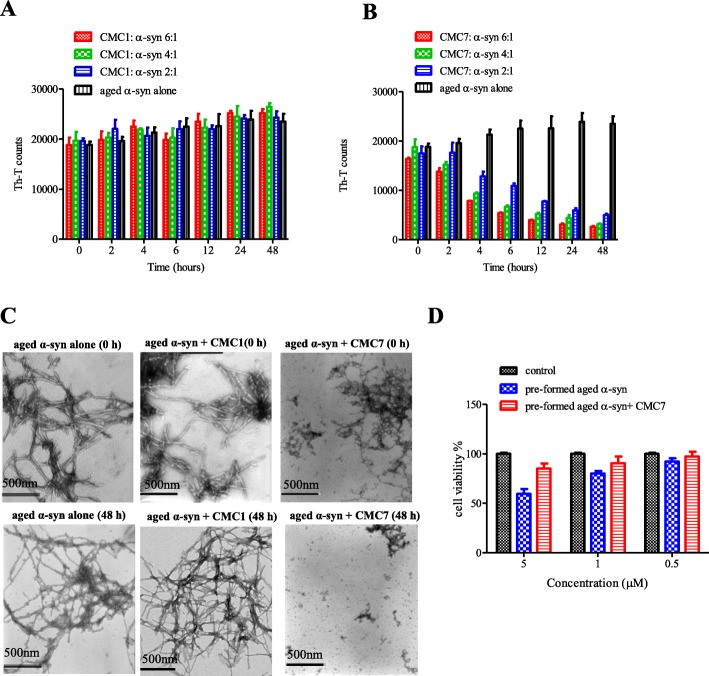
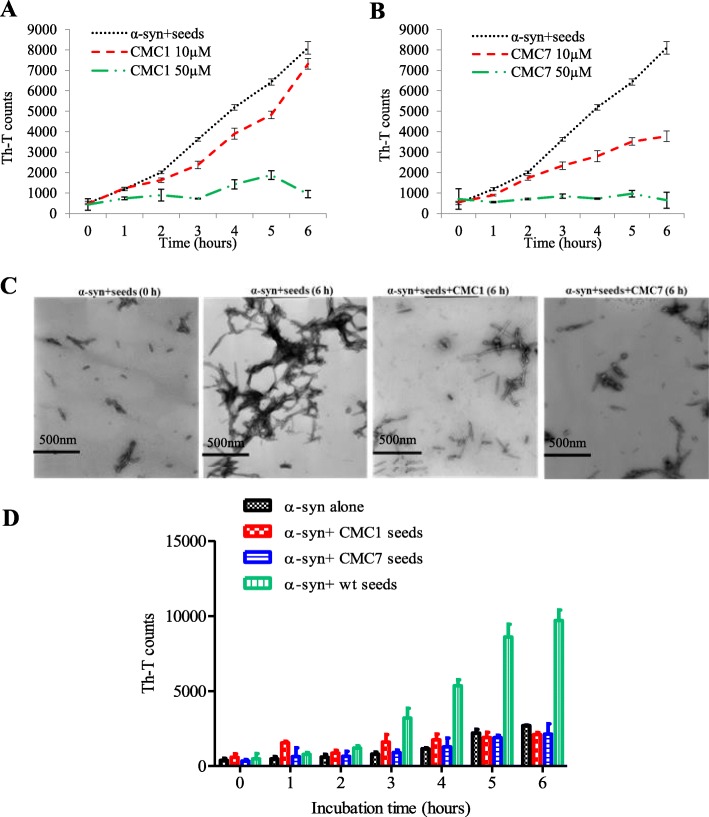
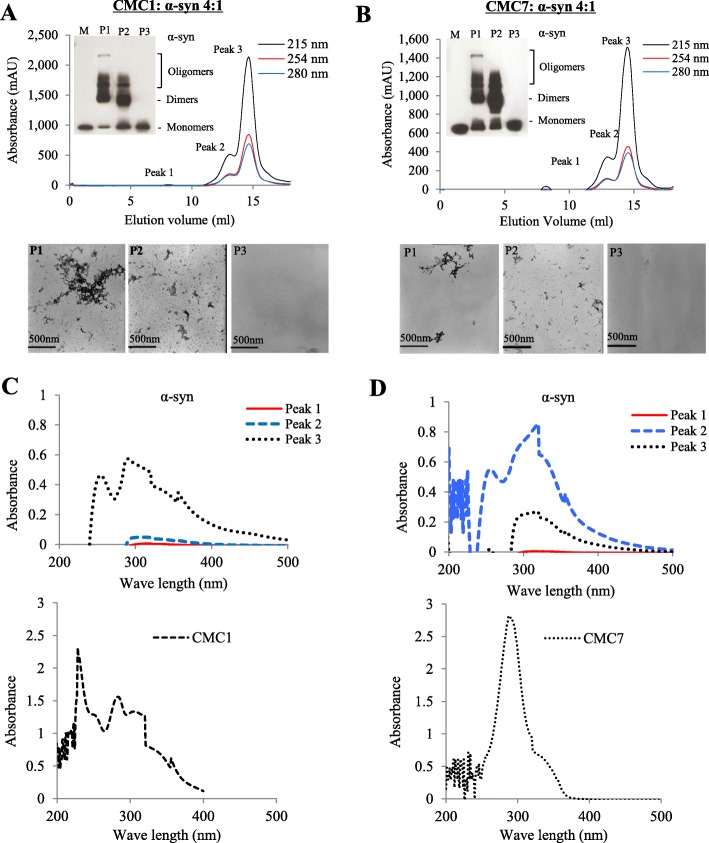
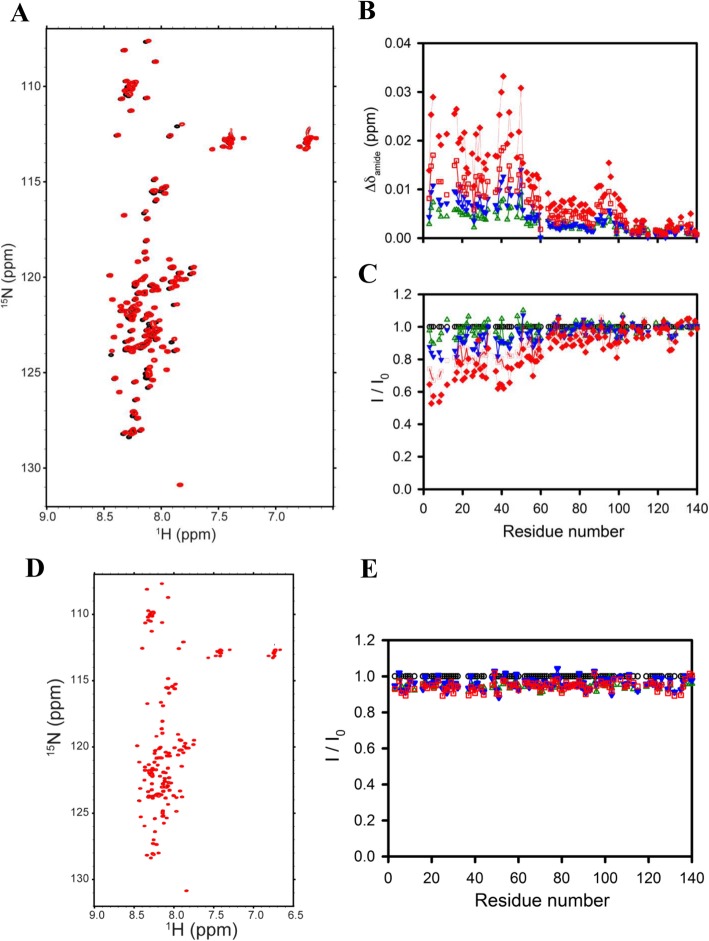
Results: Salvianolic acid B and dihydromyricetin were the two compounds that strongly inhibited the fibril growth and neurotoxicity of α-syn. In an in-vitro cell model, these compounds decreased the insoluble phosphorylated α-syn and aggregation. Also, in primary neuronal cells, these compounds showed a reduction in α-syn aggregates. Both compounds inhibited the seeded fibril growth with dihydromyricetin having the ability to disaggregate preformed α-syn fibrils. In order to investigate the inhibitory mechanisms of these two compounds towards fibril formation, we demonstrated that salvianolic acid B binds predominantly to monomers, while dihydromyricetin binds to oligomeric species and to a lower extent to monomers. Remarkably, these two compounds stabilized the soluble non-toxic oligomers lacking β-sheet content after subjecting them to proteinase K digestion.
Conclusions: Eleven compounds were tested but only two showed inhibition of α-syn aggregation, seeded fibril formation and toxicity in vitro. These findings highlight an essential beginning for development of new molecules in the field of synucleinopathies treatment.
Keywords: Amyloid fibrils; Dihydromyricetin; Parkinson’s disease; Salvianolic acid B; Seeded fibril formation; α-Synuclein.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
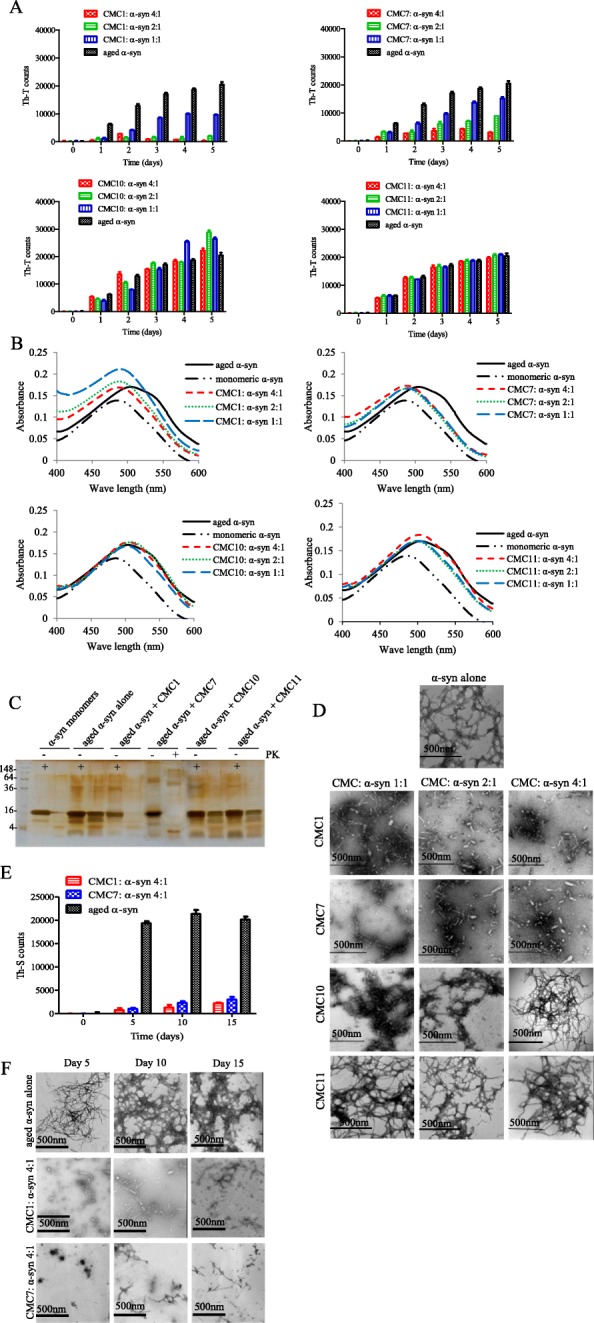
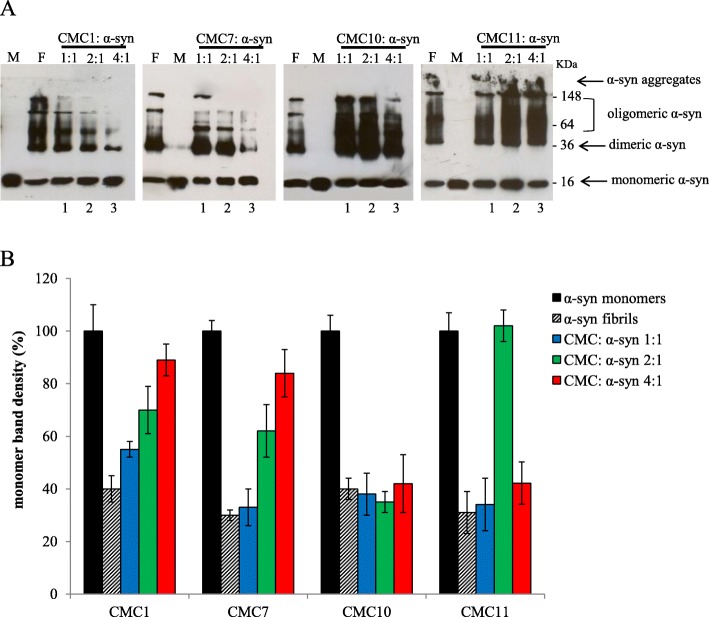
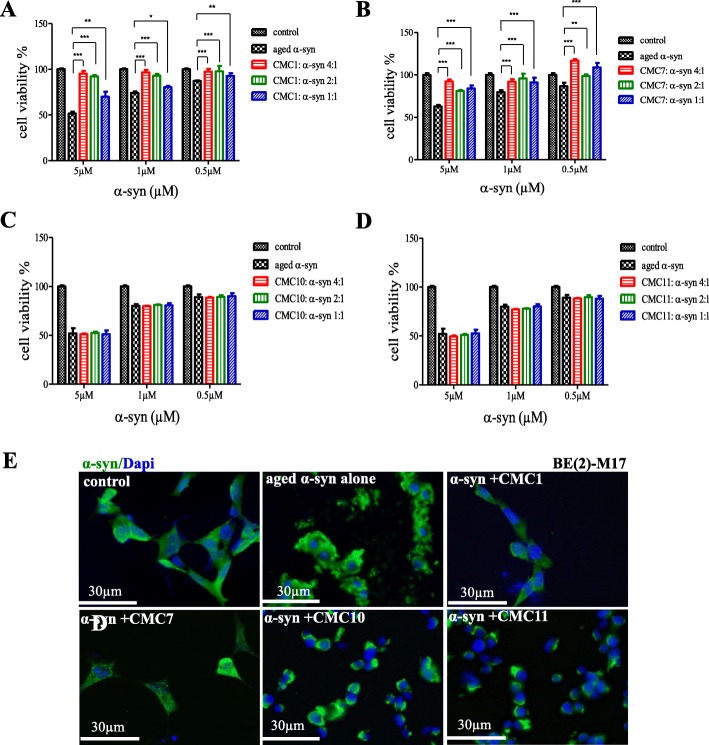
References
-
- Galvin JE, Lee VM, Schmidt ML, Tu PH, Iwatsubo T, Trojanowski JQ. Pathobiology of the Lewy body. Adv Neurol. 1999;80:313–324. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
