

Inter-α-inhibitor heavy chain-1 has an integrin-like 3D structure mediating immune regulatory activities and matrix stabilization during ovulation
- PMID: 32144206
- PMCID: PMC7170535
- DOI: 10.1074/jbc.RA119.011916
Inter-α-inhibitor heavy chain-1 has an integrin-like 3D structure mediating immune regulatory activities and matrix stabilization during ovulation
Abstract
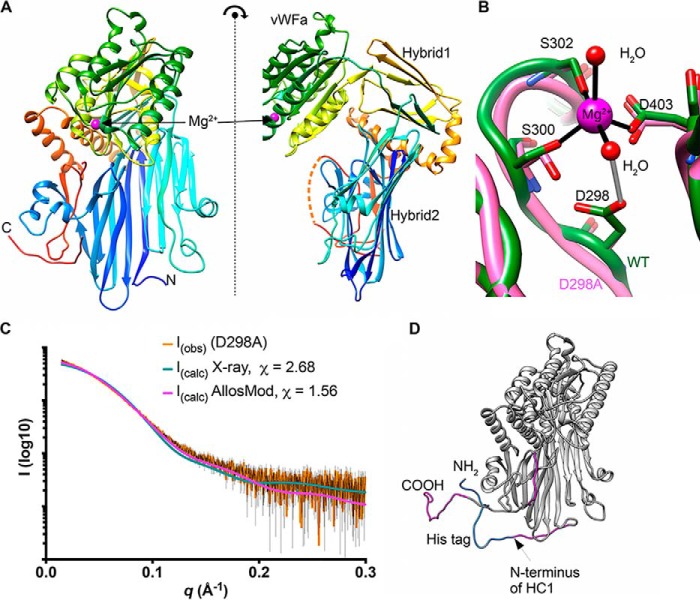
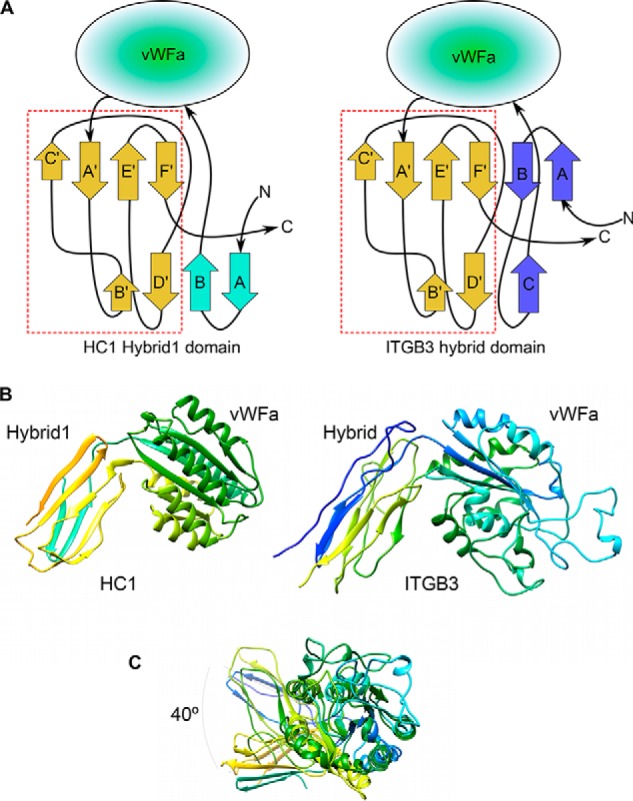
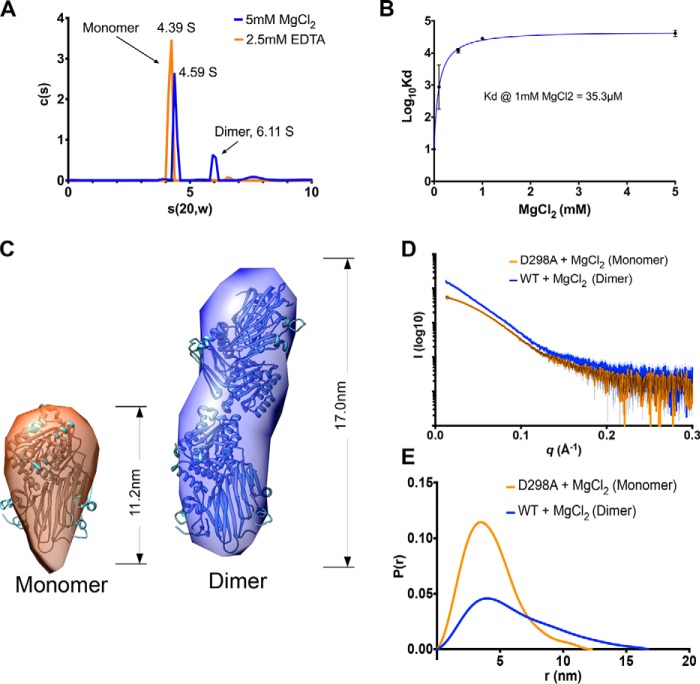
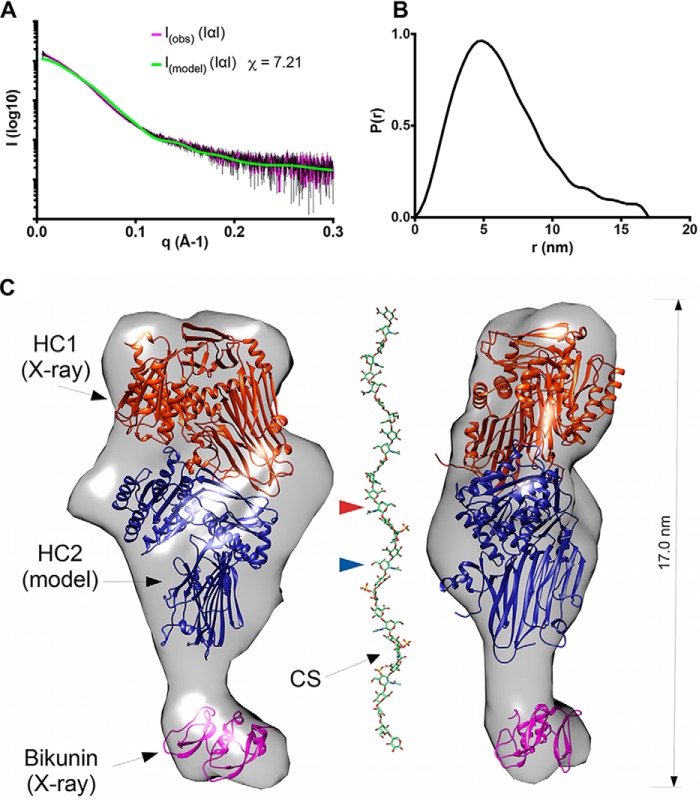
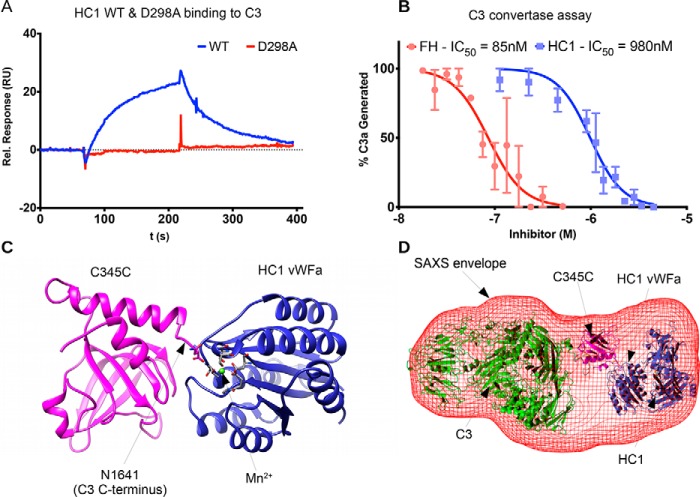
Inter-α-inhibitor is a proteoglycan essential for mammalian reproduction and also plays a less well-characterized role in inflammation. It comprises two homologous "heavy chains" (HC1 and HC2) covalently attached to chondroitin sulfate on the bikunin core protein. Before ovulation, HCs are transferred onto the polysaccharide hyaluronan (HA) to form covalent HC·HA complexes, thereby stabilizing an extracellular matrix around the oocyte required for fertilization. Additionally, such complexes form during inflammatory processes and mediate leukocyte adhesion in the synovial fluids of arthritis patients and protect against sepsis. Here using X-ray crystallography, we show that human HC1 has a structure similar to integrin β-chains, with a von Willebrand factor A domain containing a functional metal ion-dependent adhesion site (MIDAS) and an associated hybrid domain. A comparison of the WT protein and a variant with an impaired MIDAS (but otherwise structurally identical) by small-angle X-ray scattering and analytical ultracentrifugation revealed that HC1 self-associates in a cation-dependent manner, providing a mechanism for HC·HA cross-linking and matrix stabilization. Surprisingly, unlike integrins, HC1 interacted with RGD-containing ligands, such as fibronectin, vitronectin, and the latency-associated peptides of transforming growth factor β, in a MIDAS/cation-independent manner. However, HC1 utilizes its MIDAS motif to bind to and inhibit the cleavage of complement C3, and small-angle X-ray scattering-based modeling indicates that this occurs through the inhibition of the alternative pathway C3 convertase. These findings provide detailed structural and functional insights into HC1 as a regulator of innate immunity and further elucidate the role of HC·HA complexes in inflammation and ovulation.
Keywords: Inter-α-inhibitor Heavy Chain; X-ray crystallography; extracellular matrix; hyaluronan; inflammation; innate immunity; protein stability; proteoglycan; reproduction; small-angle X-ray scattering (SAXS).
© 2020 Briggs et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Enghild J. J., Thøgersen I. B., Pizzo S. V., and Salvesen G. (1989) Analysis of inter-α-trypsin inhibitor and a novel trypsin inhibitor, pre-α-trypsin inhibitor, from human plasma: polypeptide chain stoichiometry and assembly by glycan. J. Biol. Chem. 264, 15975–15981 - PubMed
-
- Zhuo L., Yoneda M., Zhao M., Yingsung W., Yoshida N., Kitagawa Y., Kawamura K., Suzuki T., and Kimata K. (2001) Defect in SHAP-hyaluronan complex causes severe female infertility: a study by inactivation of the bikunin gene in mice. J. Biol. Chem. 276, 7693–7696 10.1074/jbc.C000899200 - DOI - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
