

The Role of Transthyretin in Oligodendrocyte Development
- PMID: 32144308
- PMCID: PMC7060235
- DOI: 10.1038/s41598-020-60699-8
The Role of Transthyretin in Oligodendrocyte Development
Abstract
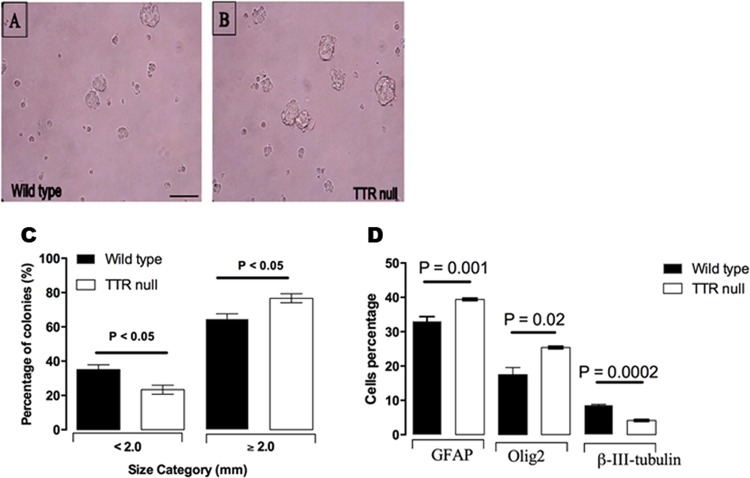
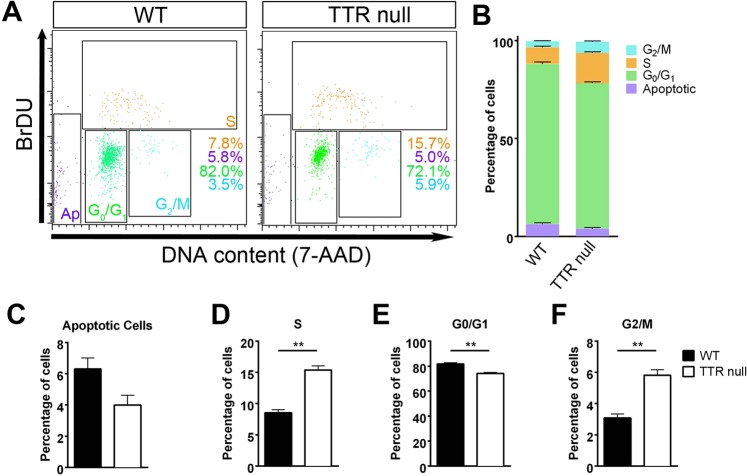
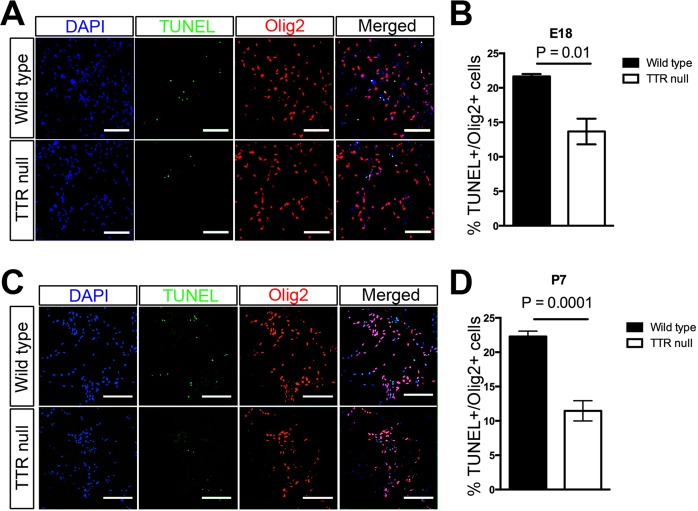
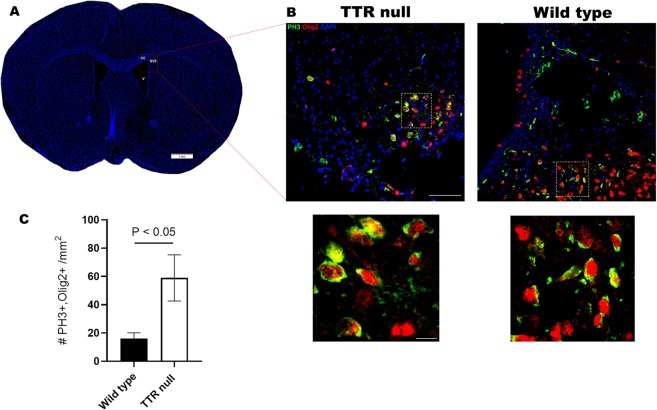
Transthyretin (TTR) is a protein that binds and distributes thyroid hormones (THs) in blood and cerebrospinal fluid. Previously, two reports identified TTR null mice as hypothyroid in the central nervous system (CNS). This prompted our investigations into developmentally regulated TH-dependent processes in brains of wildtype and TTR null mice. Despite logical expectations of a hypomyelinating phenotype in the CNS of TTR null mice, we observed a hypermyelination phenotype, synchronous with an increase in the density of oligodendrocytes in the corpus callosum and anterior commissure of TTR null mice during postnatal development. Furthermore, absence of TTR enhanced proliferation and migration of OPCs with decreased apoptosis. Neural stem cells (NSCs) isolated from the subventricular zone of TTR null mice at P21 revealed that the absence of TTR promoted NSC differentiation toward a glial lineage. Importantly, we identified TTR synthesis in OPCs, suggestive of an alternate biological function in these cells that may extend beyond an extracellular TH-distributor protein. The hypermyelination mechanism may involve increased pAKT (involved in oligodendrocyte maturation) in TTR null mice. Elucidating the regulatory role of TTR in NSC and OPC biology could lead to potential therapeutic strategies for the treatment of acquired demyelinating diseases.
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Blake CC, Geisow MJ, Swan ID, Rerat C, Rerat B. Strjcture of human plasma prealbumin at 2-5 A resolution. A preliminary report on the polypeptide chain conformation, quaternary structure and thyroxine binding. Journal of Molecular Biology. 1974;88:1–12. doi: 10.1016/0022-2836(74)90291-5. - DOI - PubMed
-
- Soprano DR, Herbert J, Soprano KJ, Schon EA, Goodman DS. Demonstration of transthyretin mRNA in the brain and other extrahepatic tissues in the rat. Journal of Biological Chemistry. 1985;260:11793–11798. - PubMed
-
- Harms PJ, et al. Transthyretin (prealbumin) gene expression in choroid plexus is strongly conserved during evolution of vertebrates. Comparative Biochemistry &. Physiology - B: Comparative Biochemistry. 1991;99:239–249. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
