

Propagating Motor Cortical Dynamics Facilitate Movement Initiation
- PMID: 32145183
- PMCID: PMC7210059
- DOI: 10.1016/j.neuron.2020.02.011
Propagating Motor Cortical Dynamics Facilitate Movement Initiation
Erratum in
-
Propagating Motor Cortical Dynamics Facilitate Movement Initiation.Neuron. 2025 Aug 6;113(15):2546. doi: 10.1016/j.neuron.2025.07.007. Epub 2025 Jul 12. Neuron. 2025. PMID: 40652940 Free PMC article. No abstract available.
Abstract
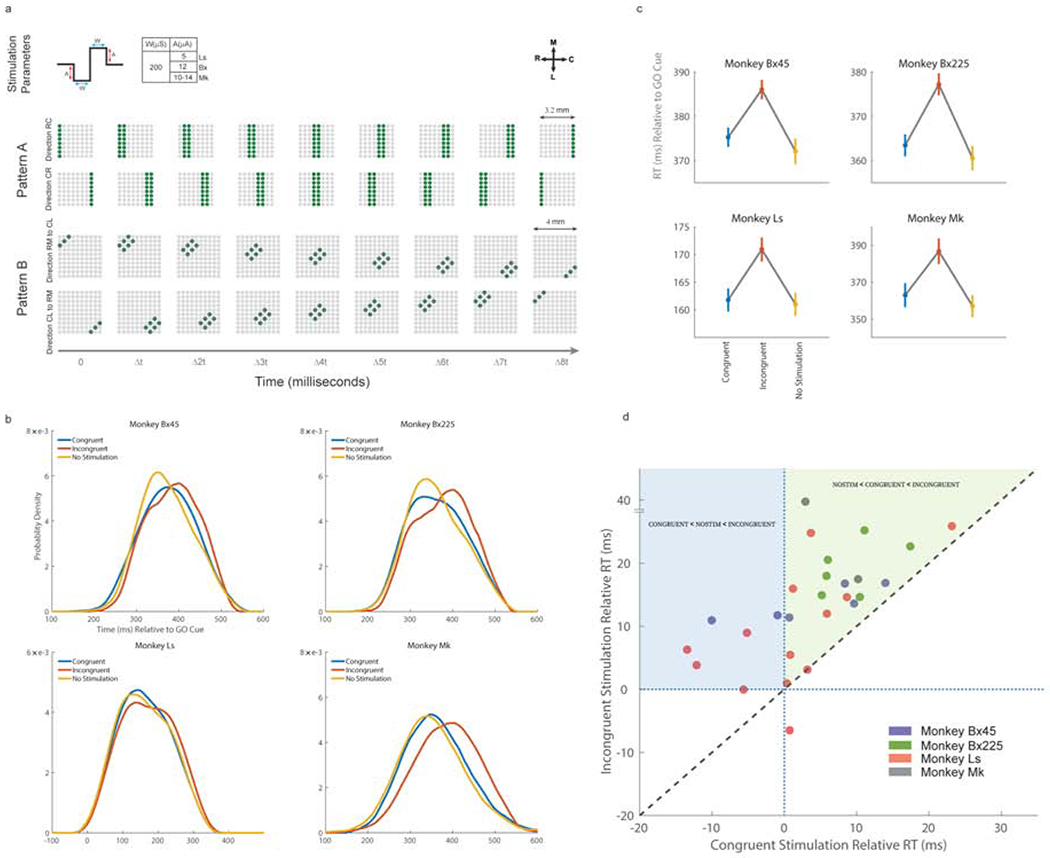
Voluntary movement initiation involves the modulations of large groups of neurons in the primary motor cortex (M1). Yet similar modulations occur during movement planning when no movement occurs. Here, we show that a sequential spatiotemporal pattern of excitability propagates across M1 prior to the movement initiation in one of two oppositely oriented directions along the rostro-caudal axis. Using spatiotemporal patterns of intracortical microstimulation, we find that reaction time increases significantly when stimulation is delivered against, but not with, the natural propagation direction. Functional connections among M1 units emerge at movement that are oriented along the same rostro-caudal axis but not during movement planning. Finally, we show that beta amplitude profiles can more accurately decode muscle activity when they conform to the natural propagating patterns. These findings provide the first causal evidence that large-scale, propagating patterns of cortical excitability are behaviorally relevant and may be a necessary component of movement initiation.
Keywords: beta attenuation; beta oscillations; functional connectivity; intracortical microstimulation; motor cortex; movement initiation; propagating patterns; spatiotemporal patterns.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests N.G.H. serves as a consultant for BlackRock Microsystems, Inc., the company that sells the multi-electrode arrays and acquisition system used in this study.
Figures





References
-
- Chase MH, 1983. Synaptic mechanisms and circuitry involved in motoneuron control during sleep. Int Rev Neurobiol 24, 213–258. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
