

Adenosine-producing regulatory B cells in head and neck cancer
- PMID: 32146518
- PMCID: PMC7303082
- DOI: 10.1007/s00262-020-02535-6
Adenosine-producing regulatory B cells in head and neck cancer
Abstract
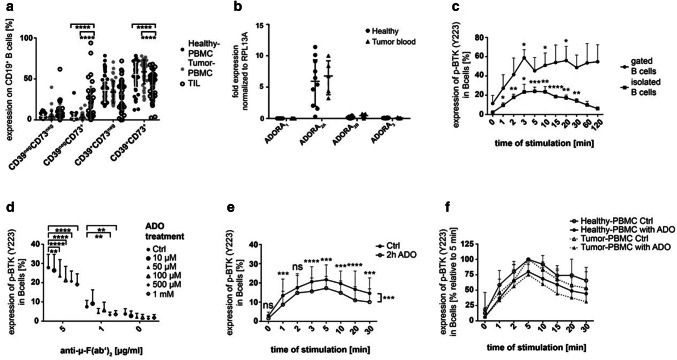
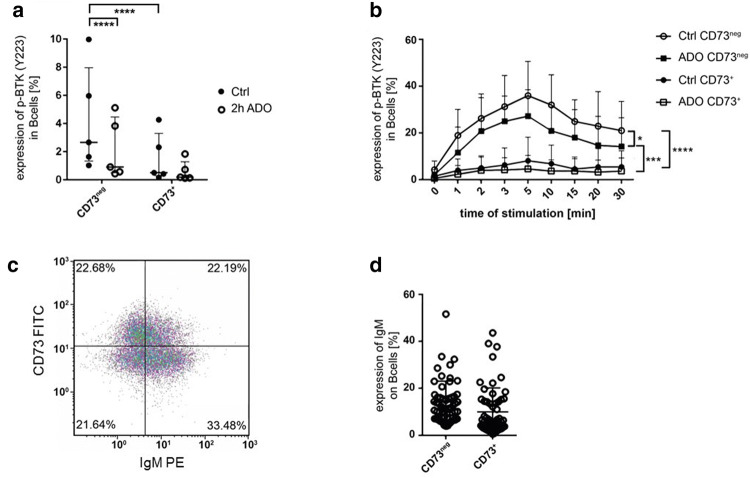
Background: Multiple mechanisms of immunosuppression have been identified in the tumor microenvironment including regulatory B cells (Breg). Recently, we have shown that Breg suppress T cell function by production of adenosine (ADO). However, the autocrine effect of ADO on B cells and the role of Breg in head and neck cancer remains unclear.
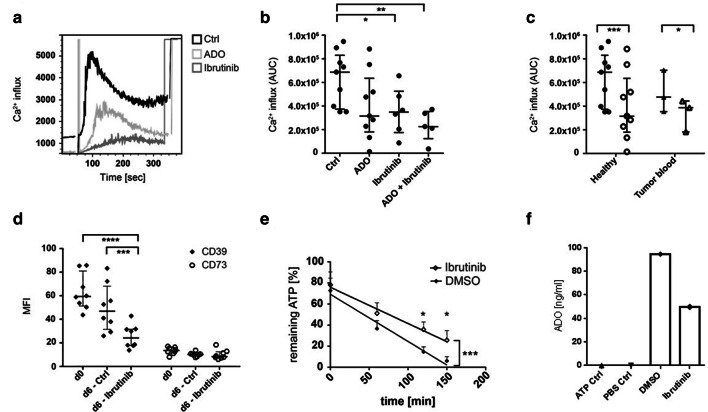
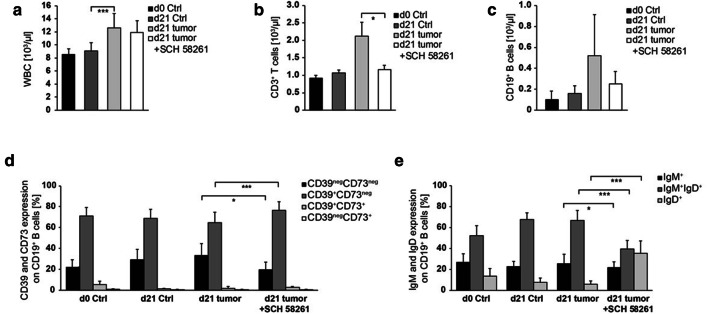
Methods: Blood (n = 42) and tumor tissue (n = 39) of head and neck cancer patients and healthy donors (n = 60) were analyzed by FACS. The effect of ADO on phenotype, intracellular signaling pathways, Ca2+ influx and ADO production was analyzed in Breg and effector B cells (Beff) by FACS, luminescence and mass spectrometry. The blockage of the ADO receptor A2A was analyzed in a murine head and neck cancer model.
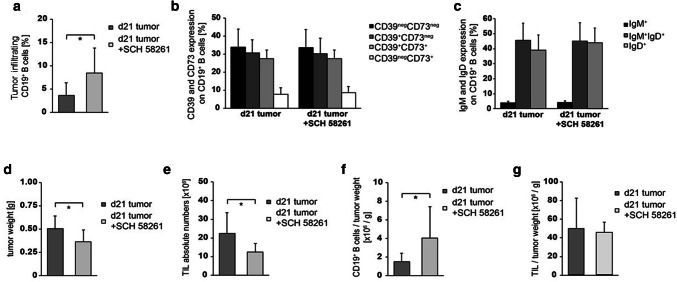
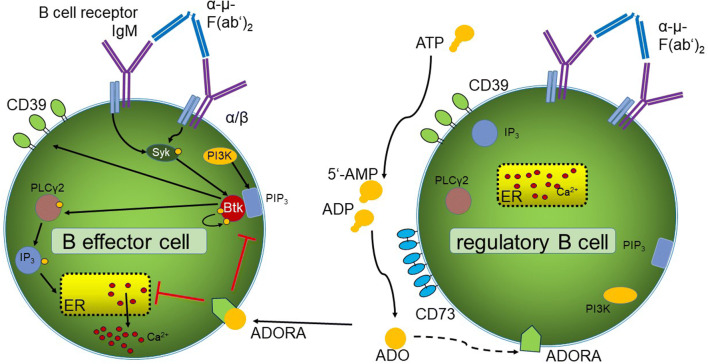
Results: ADO-producing Breg were found in tumor tissue and peripheral blood. ADO inhibited the intracellular Bruton's tyrosine kinase (BTK) and Ca2+ influx only in Beff. The inhibition of BTK by ibrutinib mimicked the effect of ADO, and ibrutinib reduced the production of ADO by downregulation of CD39 in vitro. The inhibition of ADO receptor A2A significantly reduced tumor mass and increased B cell infiltration, in vivo.
Conclusion: Our data demonstrate the presence of a novel ADO-producing Breg population within the tumor microenvironment in mice and humans. A new model is proposed on how ADO-producing Breg can influence the function of Beff cells in healthy donors and cancer patients. Thus, the modulation of the ADO pathway in B cells may serve as a therapeutic approach for cancer patients.
Keywords: ADORA2A; Adenosine; BTK; Head and neck cancer; Regulatory B cells.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Human studies were approved by the ethics committee of the University Ulm, Helmholtzstr. 20, 89081 Ulm, Germany (#255/14). The Helsinki Declaration was followed. Mice were kept in groups of 4–5 mice in individually ventilated cages with nesting material under specific pathogen-free conditions in the animal facility at the University of Ulm. All animal experiments were performed in accordance with institutional and governmental regulations in compliance with European rules (2010/63/EU) and were approved by the regional animal ethics committee Regierungspräsidium (Tübingen, Germany), Konrad-Adenauer-Str. 20, 72072 Tübingen, Germany (TVA1270).
Figures
References
-
- Gunderson AJ, Kaneda MM, Tsujikawa T, et al. Bruton tyrosine kinase-dependent immune cell cross-talk drives pancreas cancer. Cancer Discov. 2016;6:270–285. doi: 10.1158/2159-8290.cd-15-0827. - DOI - PMC - PubMed
-
- Hashimoto A, Okada H, Jiang A, Kurosaki M, Greenberg S, Clark EA, Kurosaki T. Involvement of guanosine triphosphatases and phospholipase C-gamma2 in extracellular signal-regulated kinase, c-Jun NH2-terminal kinase, and p38 mitogen-activated protein kinase activation by the B cell antigen receptor. J Exp Med. 1998;188:1287–1295. doi: 10.1084/jem.188.7.1287. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
