

Fibrosis and cancer: A strained relationship
- PMID: 32147542
- PMCID: PMC7733542
- DOI: 10.1016/j.bbcan.2020.188356
Fibrosis and cancer: A strained relationship
Abstract
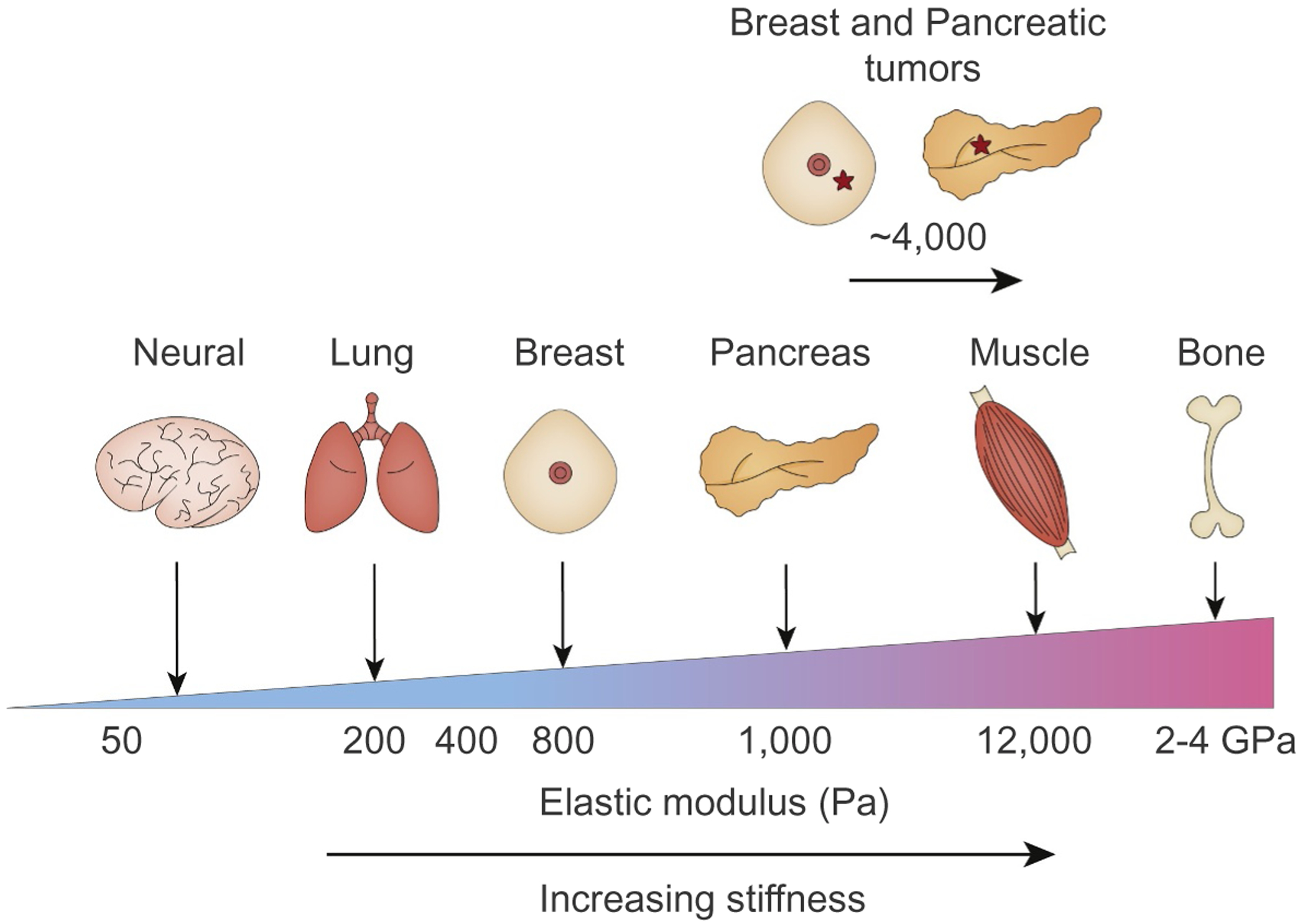
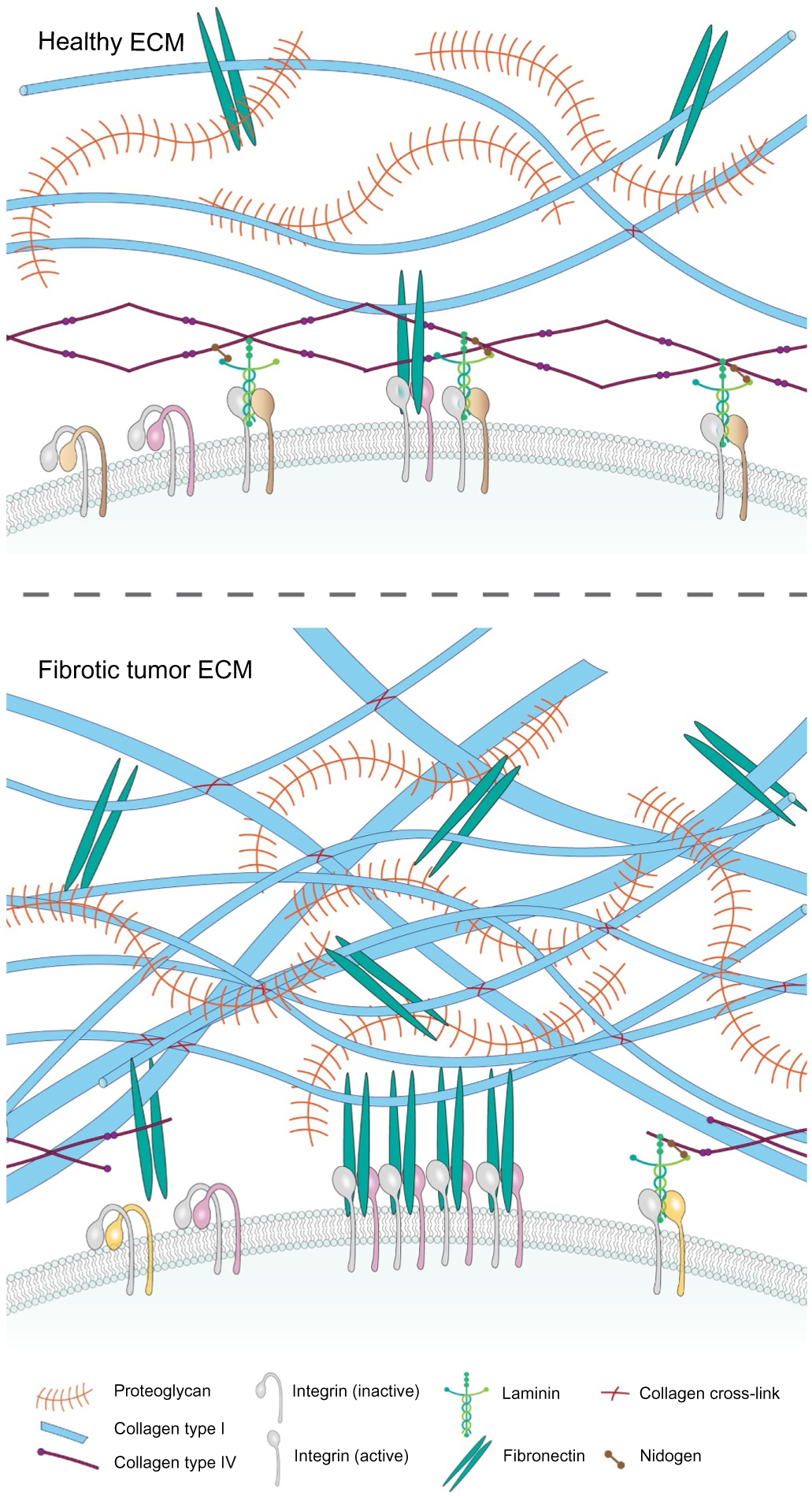
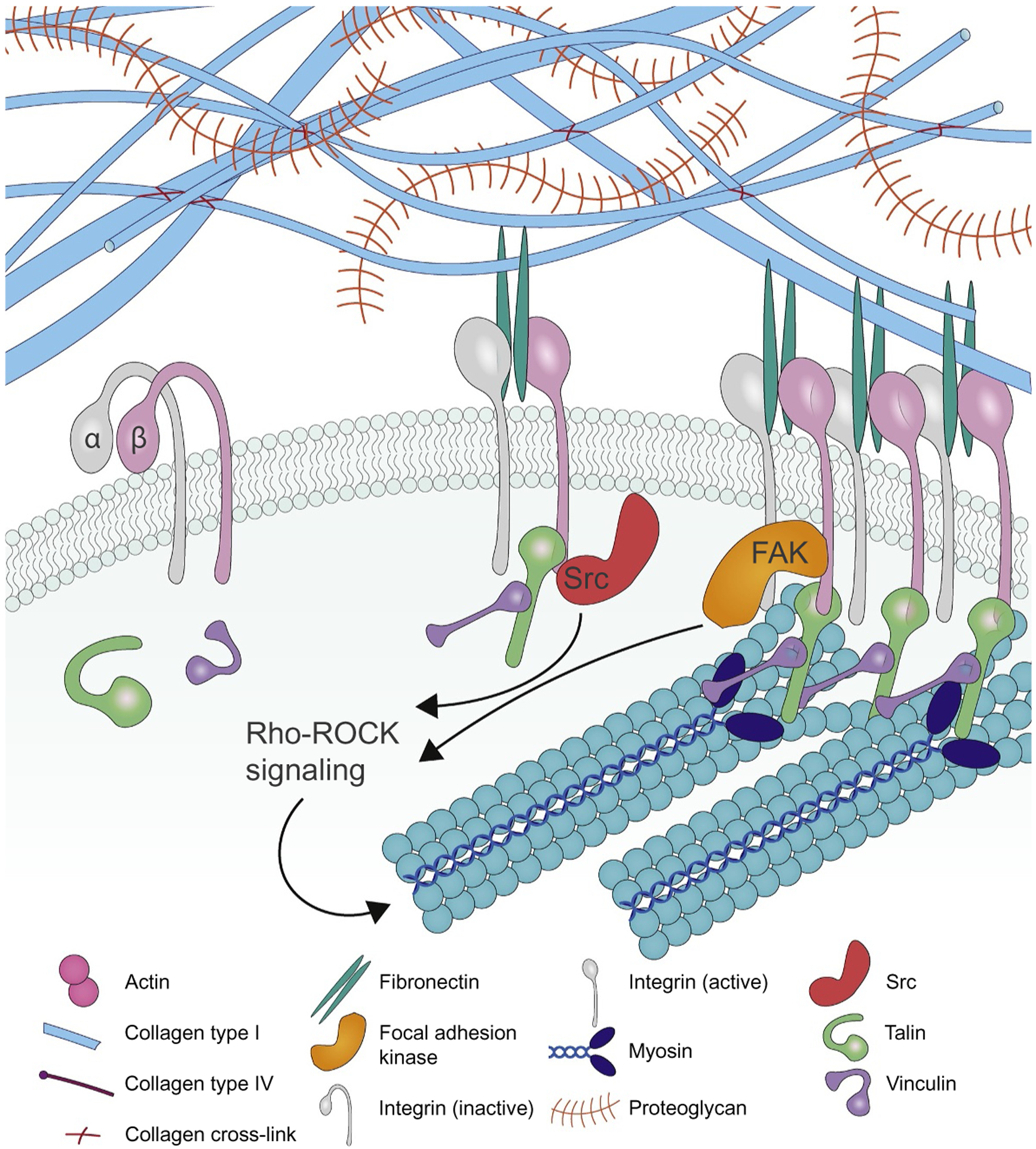
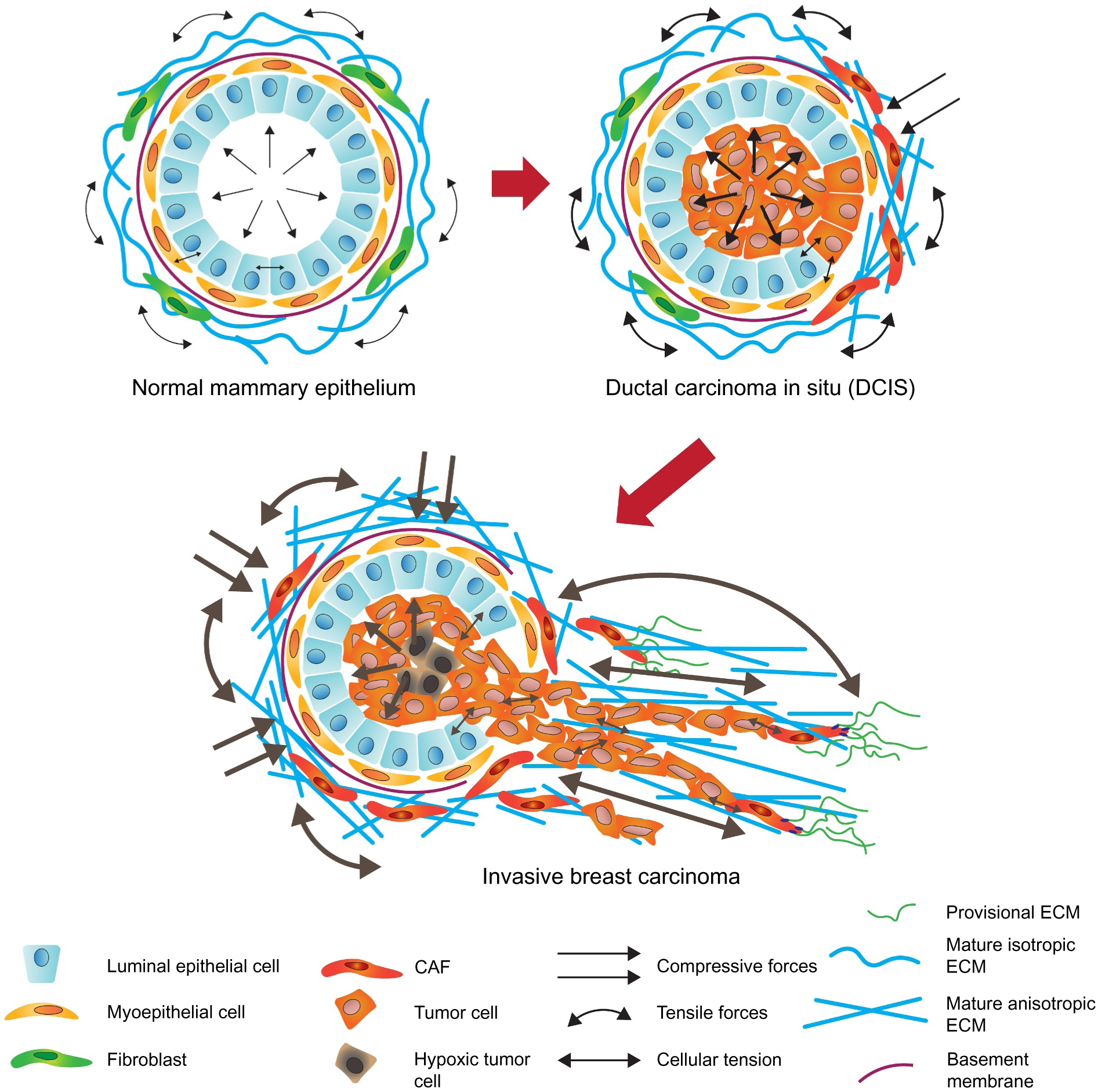
Tumors are characterized by extracellular matrix (ECM) deposition, remodeling, and cross-linking that drive fibrosis to stiffen the stroma and promote malignancy. The stiffened stroma enhances tumor cell growth, survival and migration and drives a mesenchymal transition. A stiff ECM also induces angiogenesis, hypoxia and compromises anti-tumor immunity. Not surprisingly, tumor aggression and poor patient prognosis correlate with degree of tissue fibrosis and level of stromal stiffness. In this review, we discuss the reciprocal interplay between tumor cells, cancer associated fibroblasts (CAF), immune cells and ECM stiffness in malignant transformation and cancer aggression. We discuss CAF heterogeneity and describe its impact on tumor development and aggression focusing on the role of CAFs in engineering the fibrotic tumor stroma and tuning tumor cell tension and modulating the immune response. To illustrate the role of mechanoreciprocity in tumor evolution we summarize data from breast cancer and pancreatic ductal carcinoma (PDAC) studies, and finish by discussing emerging anti-fibrotic strategies aimed at treating cancer.
Keywords: CAF; Cancer; ECM; Fibrosis; Mechanoreciprocity.
Copyright © 2020. Published by Elsevier B.V.
Conflict of interest statement
Declarations of Competing Interest
None.
Figures
References
-
- Neglia JP, et al. , The risk of cancer among patients with cystic fibrosis. Cystic fibrosis and cancer study group, N. Engl. J. Med 332 (8) (1995) 494–499. - PubMed
-
- Sørensen HT, et al. , The risk of a diagnosis of cancer after primary deep venous thrombosis or pulmonary embolism, N. Engl. J. Med 338 (17) (1998) 1169–1173. - PubMed
-
- Raimondi S, et al. , Pancreatic cancer in chronic pancreatitis; aetiology, incidence, and early detection, Best Pract. Res. Clin. Gastroenterol 24 (3) (2010) 349–358. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
