

The enigma of DNA methylation in the mammalian oocyte
- PMID: 32148772
- PMCID: PMC7043116
- DOI: 10.12688/f1000research.21513.1
The enigma of DNA methylation in the mammalian oocyte
Abstract
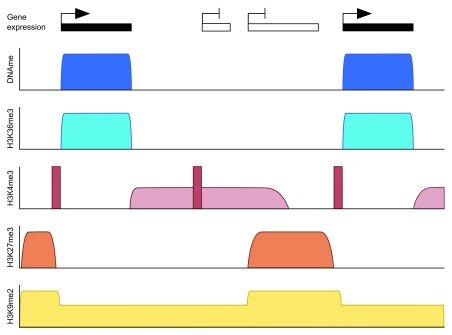
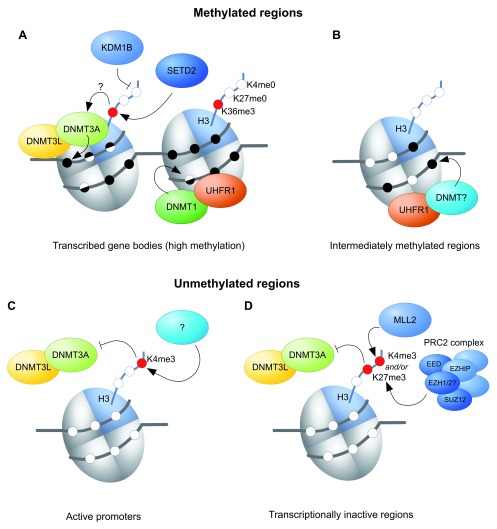
The mammalian genome experiences profound setting and resetting of epigenetic patterns during the life-course. This is understood best for DNA methylation: the specification of germ cells, gametogenesis, and early embryo development are characterised by phases of widespread erasure and rewriting of methylation. While mitigating against intergenerational transmission of epigenetic information, these processes must also ensure correct genomic imprinting that depends on faithful and long-term memory of gamete-derived methylation states in the next generation. This underscores the importance of understanding the mechanisms of methylation programming in the germline. De novo methylation in the oocyte is of particular interest because of its intimate association with transcription, which results in a bimodal methylome unique amongst mammalian cells. Moreover, this methylation landscape is entirely set up in a non-dividing cell, making the oocyte a fascinating model system in which to explore mechanistic determinants of methylation. Here, we summarise current knowledge on the oocyte DNA methylome and how it is established, focussing on recent insights from knockout models in the mouse that explore the interplay between methylation and chromatin states. We also highlight some remaining paradoxes and enigmas, in particular the involvement of non-nuclear factors for correct de novo methylation.
Keywords: DNA methylation; chromatin; oocyte.
Copyright: © 2020 Demond H and Kelsey G.
Conflict of interest statement
No competing interests were disclosed.No competing interests were disclosed.No competing interests were disclosed.No competing interests were disclosed.No competing interests were disclosed.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous