

Karyotypic Flexibility of the Complex Cancer Genome and the Role of Polyploidization in Maintenance of Structural Integrity of Cancer Chromosomes
- PMID: 32150835
- PMCID: PMC7139464
- DOI: 10.3390/cancers12030591
Karyotypic Flexibility of the Complex Cancer Genome and the Role of Polyploidization in Maintenance of Structural Integrity of Cancer Chromosomes
Abstract
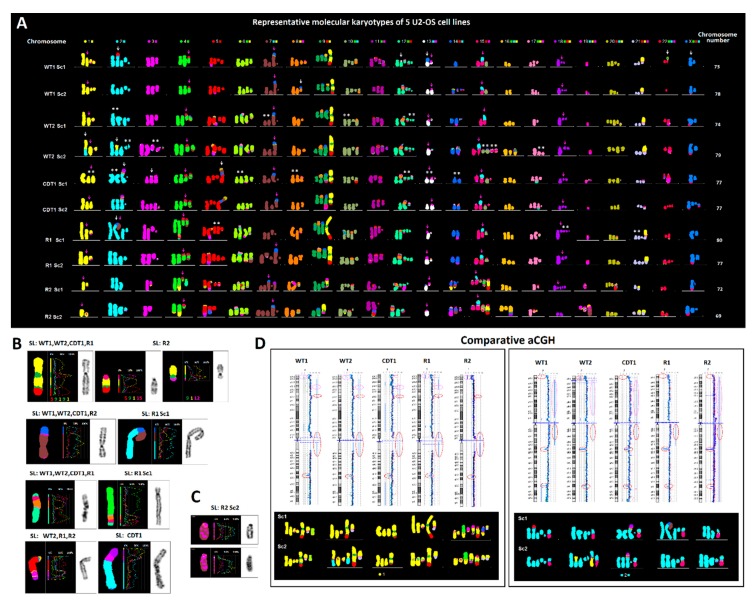
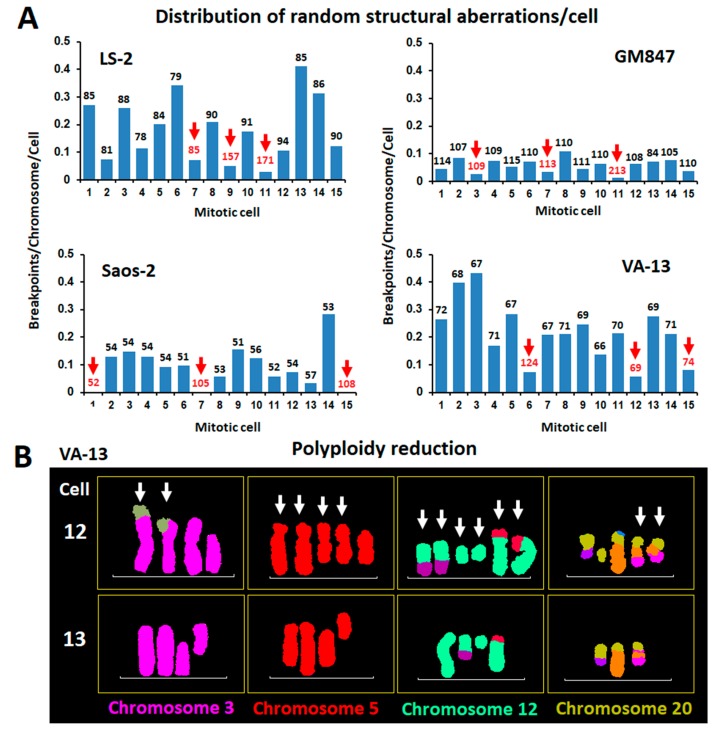
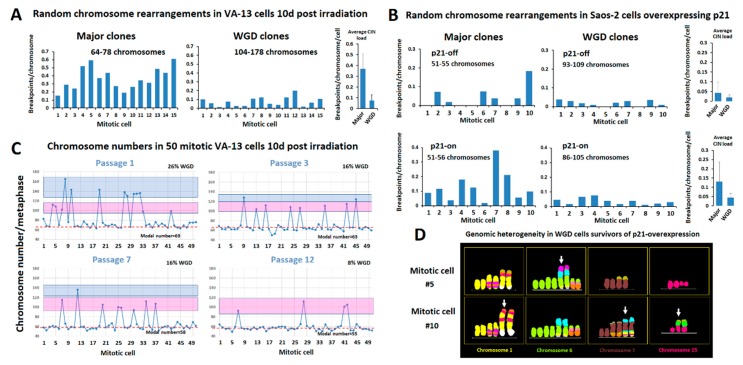
Ongoing chromosomal instability in neoplasia (CIN) generates intratumor genomic heterogeneity and limits the efficiency of oncotherapeutics. Neoplastic human cells utilizing the alternative lengthening of telomeres (ALT)-pathway, display extensive structural and numerical CIN. To unravel patterns of genome evolution driven by oncogene-replication stress, telomere dysfunction, or genotoxic therapeutic interventions, we examined by comparative genomic hybridization five karyotypically-diverse outcomes of the ALT osteosarcoma cell line U2-OS. These results demonstrate a high tendency of the complex cancer genome to perpetuate specific genomic imbalances despite the karyotypic evolution, indicating an ongoing process of genome dosage maintenance. Molecular karyotyping in four ALT human cell lines showed that mitotic cells with low levels of random structural CIN display frequent evidence of whole genome doubling (WGD), suggesting that WGD may protect clonal chromosome aberrations from hypermutation. We tested this longstanding hypothesis in ALT cells exposed to gamma irradiation or to inducible DNA replication stress under overexpression of p21. Single-cell cytogenomic analyses revealed that although polyploidization promotes genomic heterogeneity, it also protects the complex cancer genome and hence confers genotoxic therapy resistance by generating identical extra copies of driver chromosomal aberrations, which can be spared in the process of tumor evolution if they undergo unstable or unfit rearrangements.
Keywords: DNA replication stress; alternative lengthening of telomeres (ALT); chromosomal instability in neoplasia (CIN); intratumor genomic heterogeneity; polyploidy; therapy resistance; whole genome doubling (WGD).
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
LinkOut - more resources
Full Text Sources
