

Transcriptional regulation of DC fate specification
- PMID: 32151907
- PMCID: PMC7187805
- DOI: 10.1016/j.molimm.2020.02.021
Transcriptional regulation of DC fate specification
Abstract
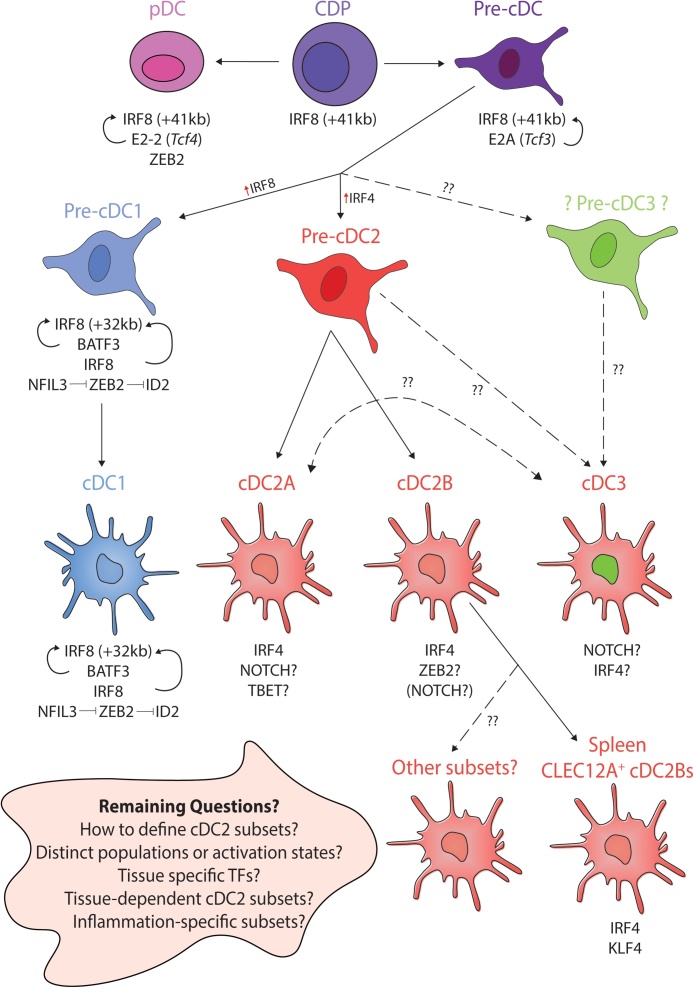
Dendritic cells function in the immune system to instruct adaptive immune cells to respond accordingly to different threats. While conventional dendritic cells can be subdivided into two main subtypes, termed cDC1s and cDC2s, it is clear that further heterogeneity exists within these subtypes, particularly for cDC2s. Understanding the signals involved in specifying each of these lineages and subtypes thereof is crucial to (i) enable us to determine their specific functions and (ii) put us in a position to be able to target these cells to promote or prevent a specific function in any given disease setting. Although we still have much to learn regarding the specification of these cells, here we review the most recent advances in our understanding of this and highlight some of the next questions for the future.
Keywords: DC fate; Dendritic cells; Irf4; Irf8; Klf4; Notch; Tbet; Transcription Factors; Zeb1; Zeb2.
Copyright © 2020 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
References
-
- Askenase M.H., Han S.-J., Byrd A.L., Morais da Fonseca D., Bouladoux N., Wilhelm C., Konkel J.E., Hand T.W., Lacerda-Queiroz N., Su X., Trinchieri G., Grainger J.R., Belkaid Y. Bone-Marrow-Resident nk cells prime monocytes for regulatory function during infection. Immunity. 2015;42:1130–1142. doi: 10.1016/j.immuni.2015.05.011. - DOI - PMC - PubMed
-
- Auderset F., Schuster S., Fasnacht N., Coutaz M., Charmoy M., Koch U., Favre S., Wilson A., Trottein F., Alexander J., Luther S.A., MacDonald H.R., Radtke F., Tacchini-Cottier F. Notch signaling regulates follicular helper t cell differentiation. J. Immunol. 2013;191:2344–2350. doi: 10.4049/jimmunol.1300643. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
