

Toward an Evolutionarily Appropriate Null Model: Jointly Inferring Demography and Purifying Selection
- PMID: 32152045
- PMCID: PMC7198275
- DOI: 10.1534/genetics.119.303002
Toward an Evolutionarily Appropriate Null Model: Jointly Inferring Demography and Purifying Selection
Abstract
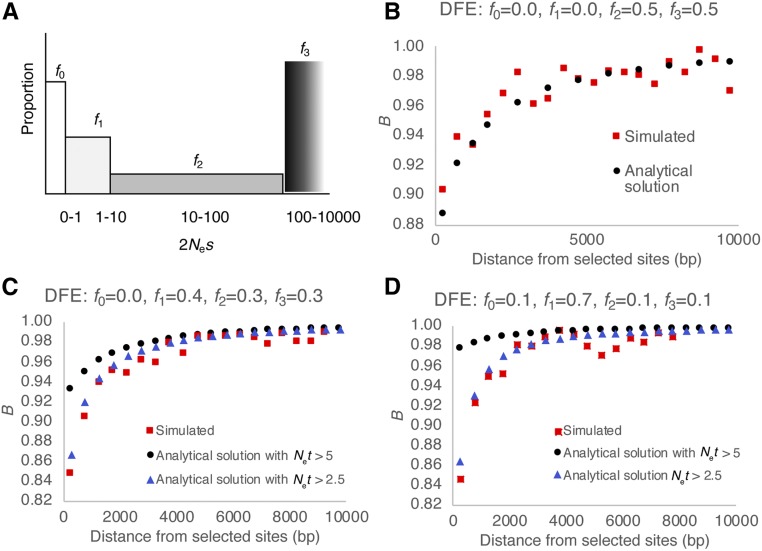
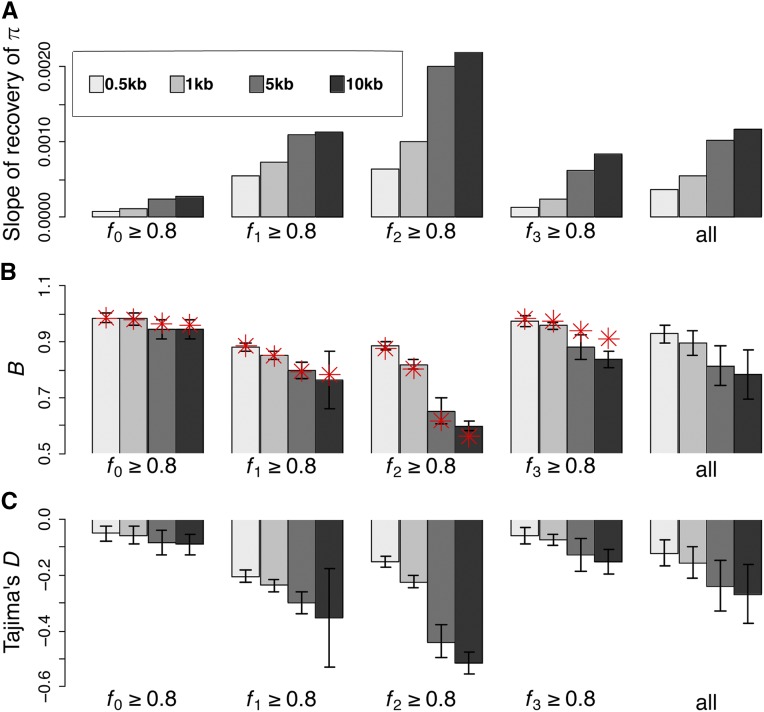
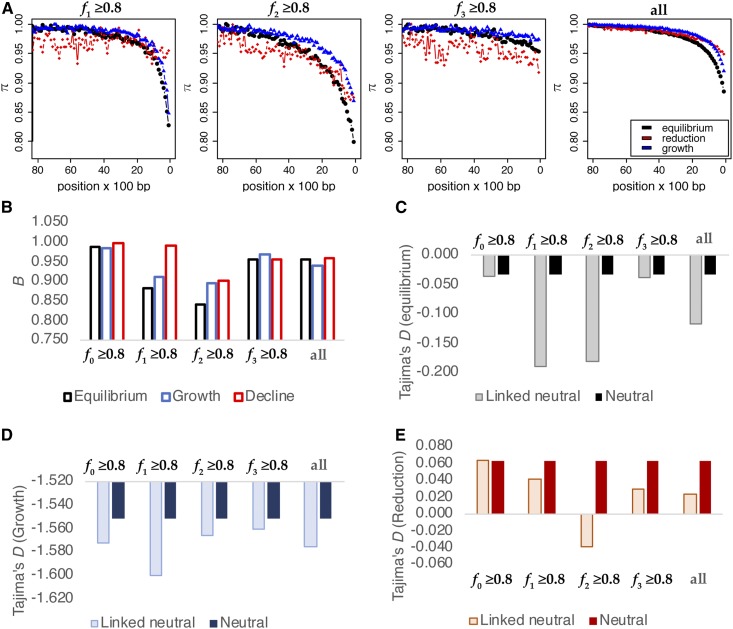
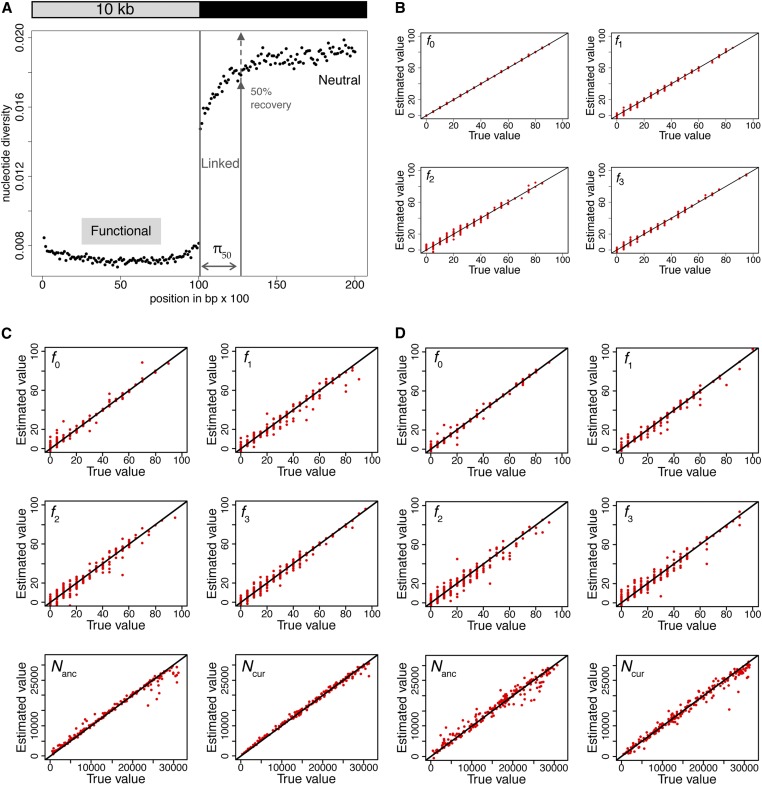
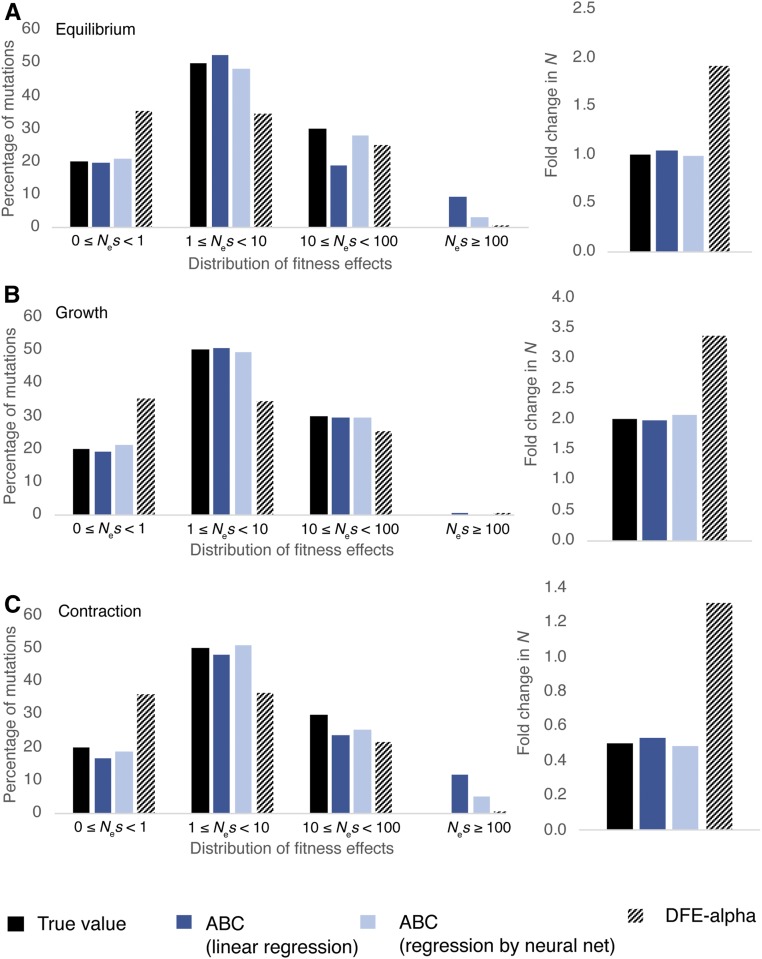
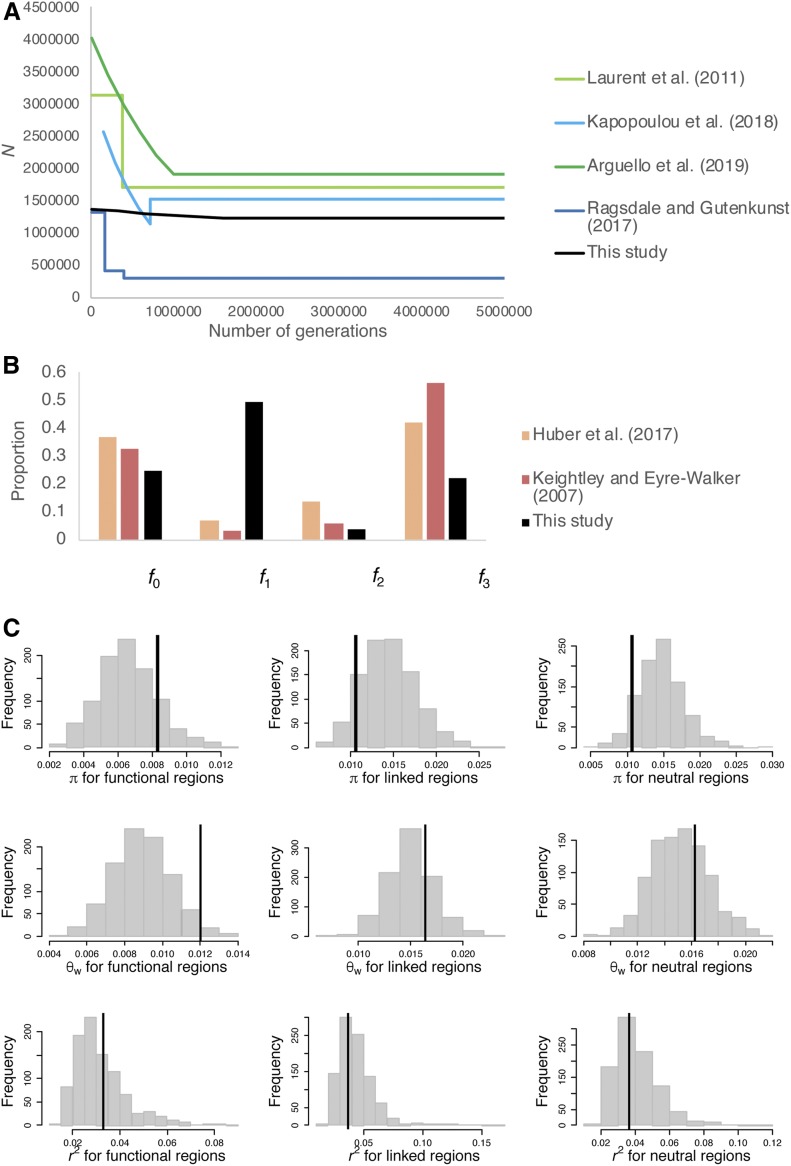
The question of the relative evolutionary roles of adaptive and nonadaptive processes has been a central debate in population genetics for nearly a century. While advances have been made in the theoretical development of the underlying models, and statistical methods for estimating their parameters from large-scale genomic data, a framework for an appropriate null model remains elusive. A model incorporating evolutionary processes known to be in constant operation, genetic drift (as modulated by the demographic history of the population) and purifying selection, is lacking. Without such a null model, the role of adaptive processes in shaping within- and between-population variation may not be accurately assessed. Here, we investigate how population size changes and the strength of purifying selection affect patterns of variation at "neutral" sites near functional genomic components. We propose a novel statistical framework for jointly inferring the contribution of the relevant selective and demographic parameters. By means of extensive performance analyses, we quantify the utility of the approach, identify the most important statistics for parameter estimation, and compare the results with existing methods. Finally, we reanalyze genome-wide population-level data from a Zambian population of Drosophila melanogaster, and find that it has experienced a much slower rate of population growth than was inferred when the effects of purifying selection were neglected. Our approach represents an appropriate null model, against which the effects of positive selection can be assessed.
Keywords: approximate Bayesian computation; background selection; demographic inference; distribution of fitness effects.
Copyright © 2020 by the Genetics Society of America.
Figures
References
-
- Bank C., Hietpas R. T., Wong A., Bolon D. N., and Jensen J. D., 2014a A Bayesian MCMC approach to assess the complete distribution of fitness effects of new mutations: uncovering the potential for adaptive walks in challenging environments. Genetics 196: 841–852. 10.1534/genetics.113.156190 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
