

Mild depolarization of the inner mitochondrial membrane is a crucial component of an anti-aging program
- PMID: 32152094
- PMCID: PMC7104298
- DOI: 10.1073/pnas.1916414117
Mild depolarization of the inner mitochondrial membrane is a crucial component of an anti-aging program
Abstract
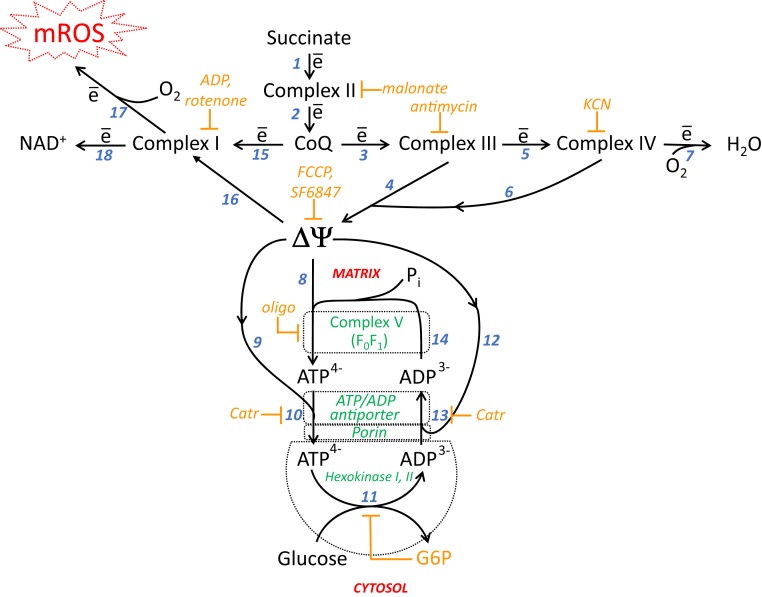
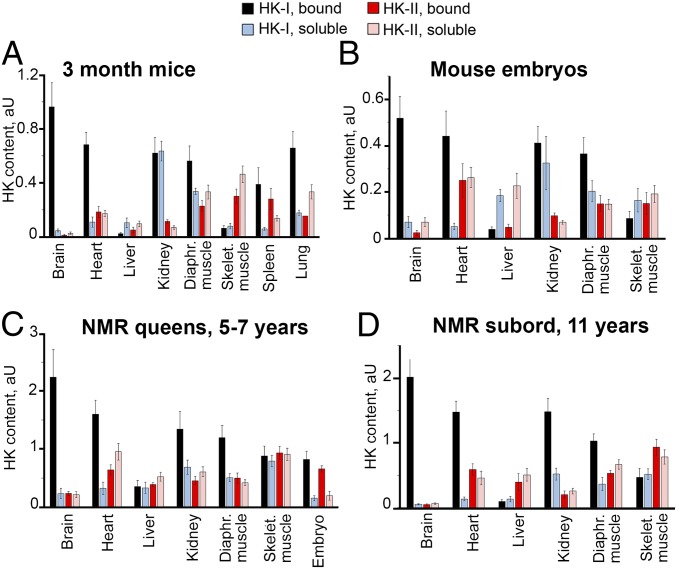
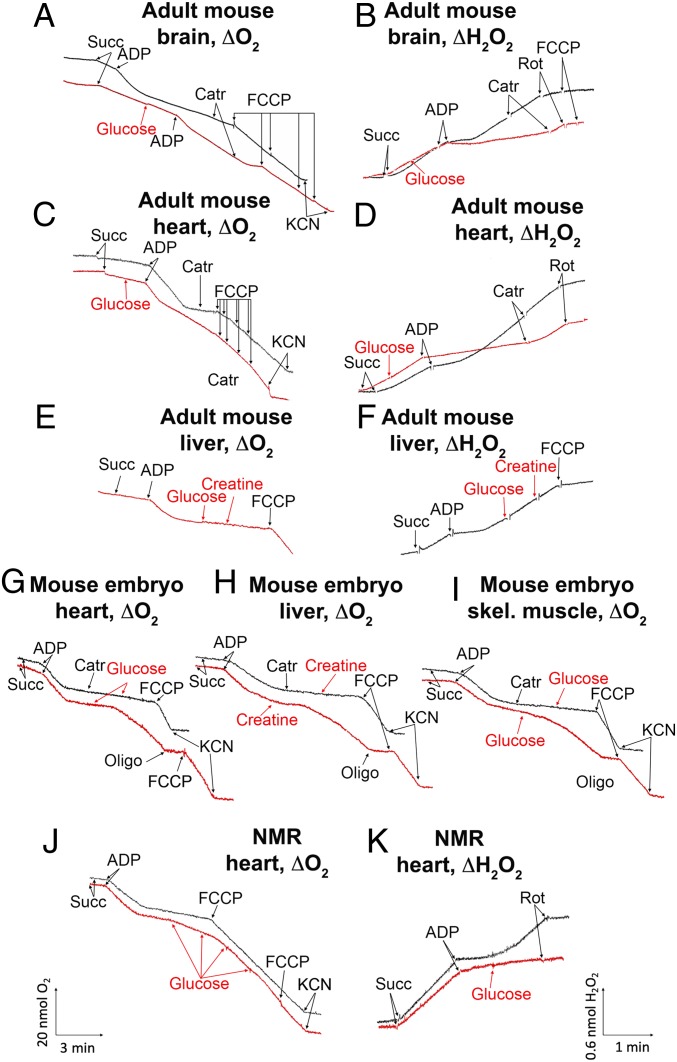
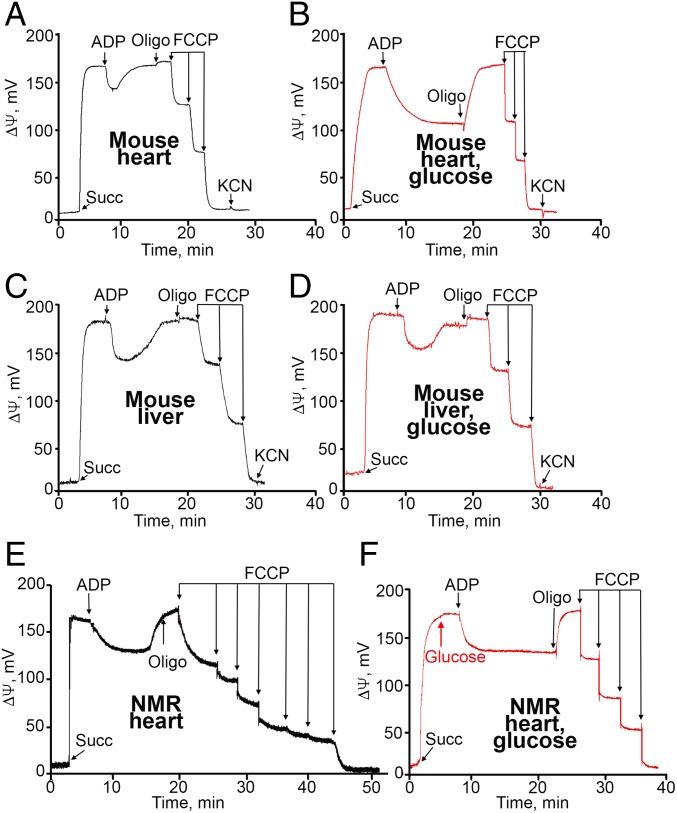
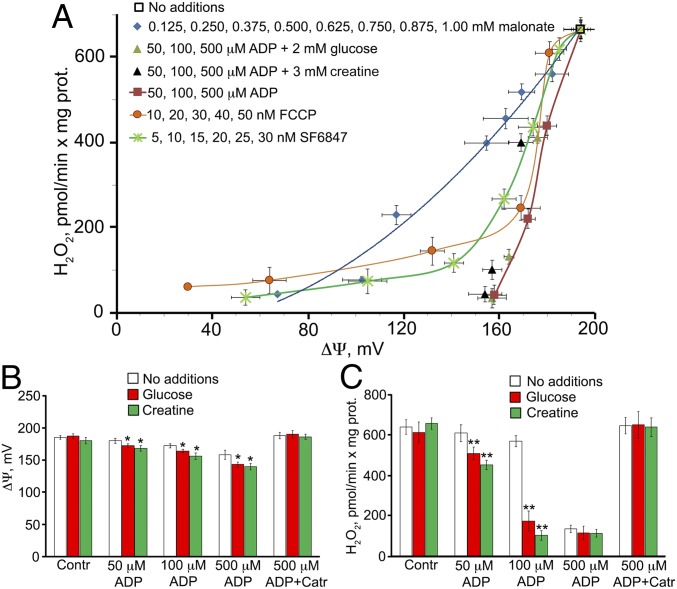
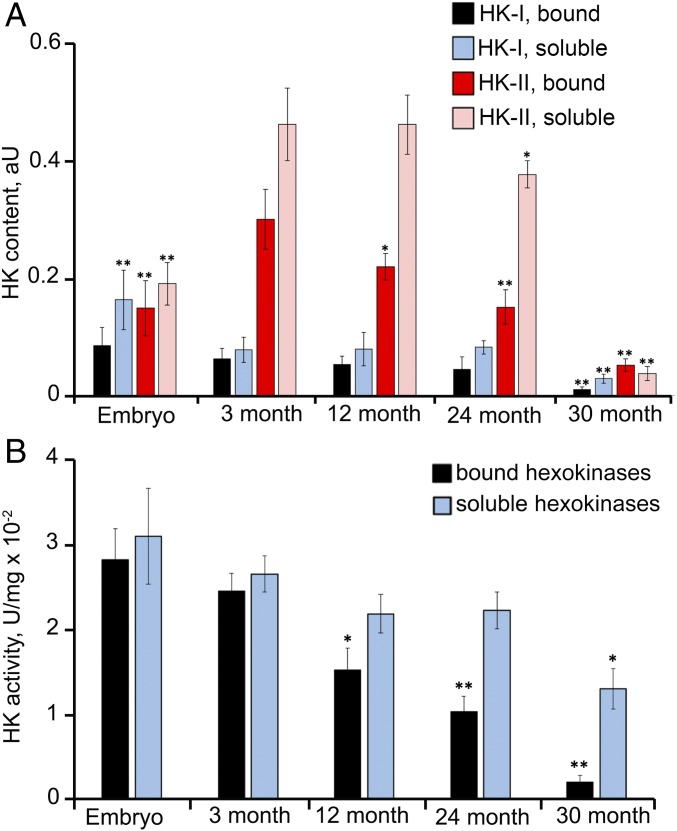
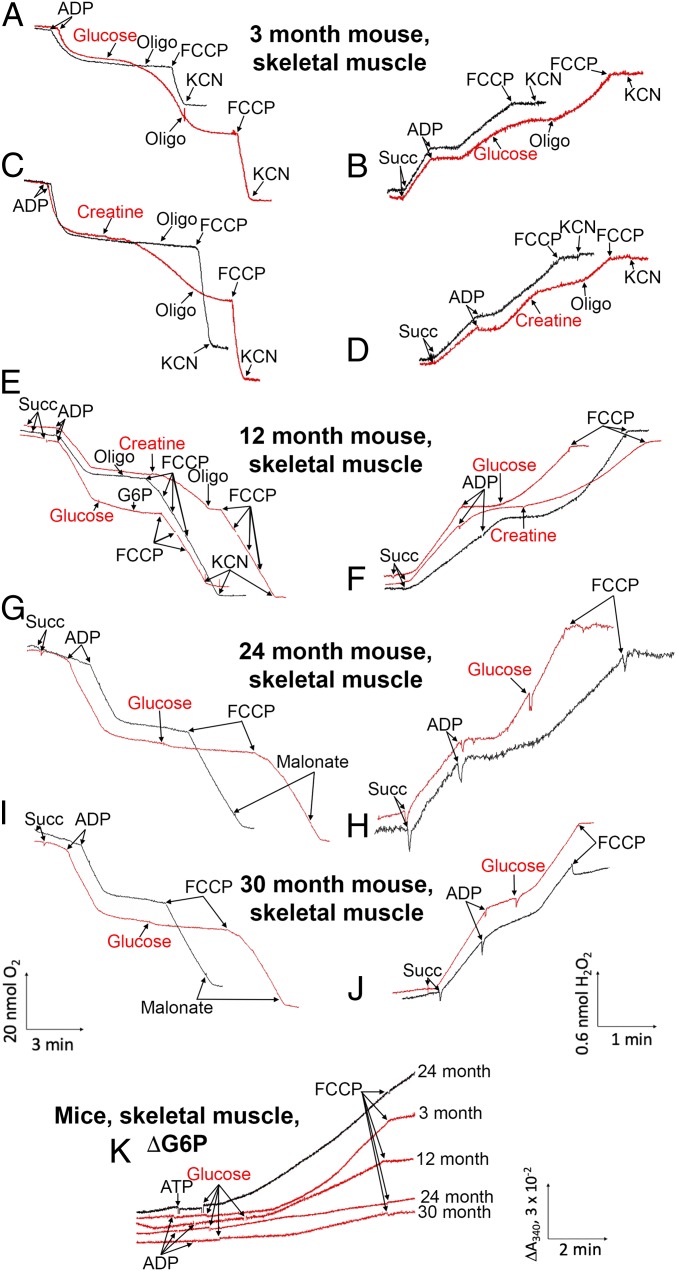
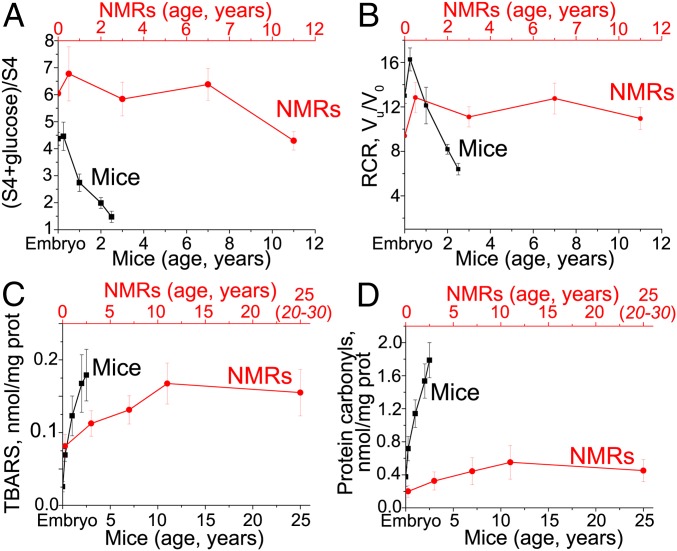
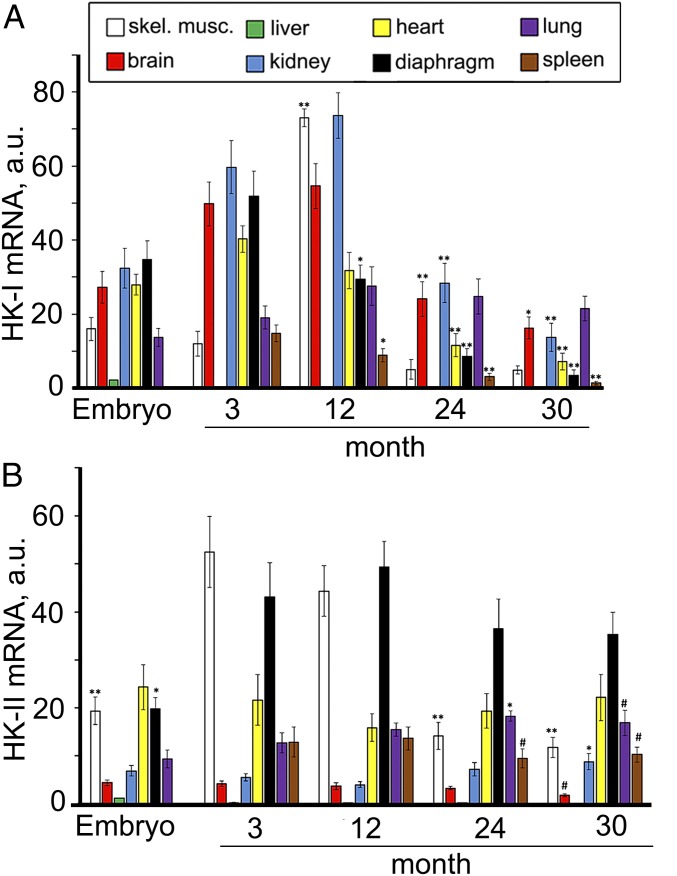
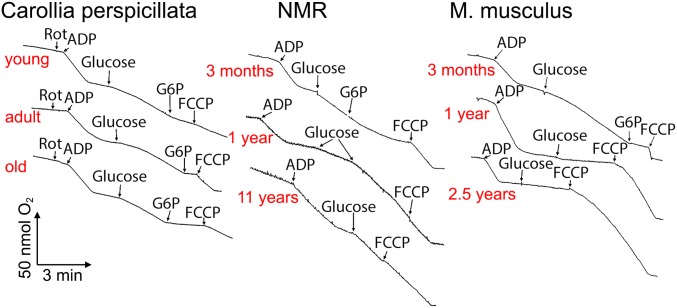
The mitochondria of various tissues from mice, naked mole rats (NMRs), and bats possess two mechanistically similar systems to prevent the generation of mitochondrial reactive oxygen species (mROS): hexokinases I and II and creatine kinase bound to mitochondrial membranes. Both systems operate in a manner such that one of the kinase substrates (mitochondrial ATP) is electrophoretically transported by the ATP/ADP antiporter to the catalytic site of bound hexokinase or bound creatine kinase without ATP dilution in the cytosol. One of the kinase reaction products, ADP, is transported back to the mitochondrial matrix via the antiporter, again through an electrophoretic process without cytosol dilution. The system in question continuously supports H+-ATP synthase with ADP until glucose or creatine is available. Under these conditions, the membrane potential, ∆ψ, is maintained at a lower than maximal level (i.e., mild depolarization of mitochondria). This ∆ψ decrease is sufficient to completely inhibit mROS generation. In 2.5-y-old mice, mild depolarization disappears in the skeletal muscles, diaphragm, heart, spleen, and brain and partially in the lung and kidney. This age-dependent decrease in the levels of bound kinases is not observed in NMRs and bats for many years. As a result, ROS-mediated protein damage, which is substantial during the aging of short-lived mice, is stabilized at low levels during the aging of long-lived NMRs and bats. It is suggested that this mitochondrial mild depolarization is a crucial component of the mitochondrial anti-aging system.
Keywords: aging; antioxidant; mild depolarization; mitochondria; naked mole rat.
Copyright © 2020 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
VDAC electronics: 2. A new, anaerobic mechanism of generation of the membrane potentials in mitochondria.Biochim Biophys Acta. 2014 Jul;1838(7):1801-8. doi: 10.1016/j.bbamem.2014.02.007. Epub 2014 Feb 22. Biochim Biophys Acta. 2014. PMID: 24565793
-
Mitochondrial creatine kinase activity prevents reactive oxygen species generation: antioxidant role of mitochondrial kinase-dependent ADP re-cycling activity.J Biol Chem. 2006 Dec 8;281(49):37361-71. doi: 10.1074/jbc.M604123200. Epub 2006 Oct 6. J Biol Chem. 2006. PMID: 17028195
-
Mitochondria and diabetes. Genetic, biochemical, and clinical implications of the cellular energy circuit.Diabetes. 1996 Feb;45(2):113-26. doi: 10.2337/diab.45.2.113. Diabetes. 1996. PMID: 8549853 Review.
-
Apparent "mild depolarization of the inner mitochondrial membrane" as a result of a possible generation of the outer membrane potential.Biochim Biophys Acta Biomembr. 2022 Nov 1;1864(11):184032. doi: 10.1016/j.bbamem.2022.184032. Epub 2022 Aug 16. Biochim Biophys Acta Biomembr. 2022. PMID: 35985076
-
The mitochondrial energy transduction system and the aging process.Am J Physiol Cell Physiol. 2007 Feb;292(2):C670-86. doi: 10.1152/ajpcell.00213.2006. Epub 2006 Oct 4. Am J Physiol Cell Physiol. 2007. PMID: 17020935 Review.
Cited by
-
BNIP3 as a new tool to promote healthy brain aging.Aging Cell. 2024 Feb;23(2):e14042. doi: 10.1111/acel.14042. Epub 2023 Nov 29. Aging Cell. 2024. PMID: 38030595 Free PMC article. Review.
-
OMA1 protease eliminates arrested protein import intermediates upon mitochondrial depolarization.J Cell Biol. 2024 May 6;223(5):e202306051. doi: 10.1083/jcb.202306051. Epub 2024 Mar 26. J Cell Biol. 2024. PMID: 38530280 Free PMC article.
-
Aging and aging-related diseases: from molecular mechanisms to interventions and treatments.Signal Transduct Target Ther. 2022 Dec 16;7(1):391. doi: 10.1038/s41392-022-01251-0. Signal Transduct Target Ther. 2022. PMID: 36522308 Free PMC article. Review.
-
An anti-glioblastoma gold(i)-NHC complex distorts mitochondrial morphology and bioenergetics to induce tumor growth inhibition.RSC Chem Biol. 2023 May 25;4(8):592-599. doi: 10.1039/d3cb00051f. eCollection 2023 Aug 3. RSC Chem Biol. 2023. PMID: 37547458 Free PMC article.
-
Mitochondria and telomeres: hand in glove.Biogerontology. 2024 Apr;25(2):289-300. doi: 10.1007/s10522-023-10074-7. Epub 2023 Oct 21. Biogerontology. 2024. PMID: 37864609 Review.
References
-
- Korshunov S. S., Skulachev V. P., Starkov A. A., High protonic potential actuates a mechanism of production of reactive oxygen species in mitochondria. FEBS Lett. 416, 15–18 (1997). - PubMed
-
- da-Silva W. S., et al. , Mitochondrial bound hexokinase activity as a preventive antioxidant defense: Steady-state ADP formation as a regulatory mechanism of membrane potential and reactive oxygen species generation in mitochondria. J. Biol. Chem. 279, 39846–39855 (2004). - PubMed
-
- Meyer L. E., et al. , Mitochondrial creatine kinase activity prevents reactive oxygen species generation: Antioxidant role of mitochondrial kinase-dependent ADP re-cycling activity. J. Biol. Chem. 281, 37361–37371 (2006). - PubMed
-
- Santiago A. P., Chaves E. A., Oliveira M. F., Galina A., Reactive oxygen species generation is modulated by mitochondrial kinases: Correlation with mitochondrial antioxidant peroxidases in rat tissues. Biochimie 90, 1566–1577 (2008). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
