

Deep mutational scanning reveals the structural basis for α-synuclein activity
- PMID: 32152544
- PMCID: PMC7339969
- DOI: 10.1038/s41589-020-0480-6
Deep mutational scanning reveals the structural basis for α-synuclein activity
Abstract
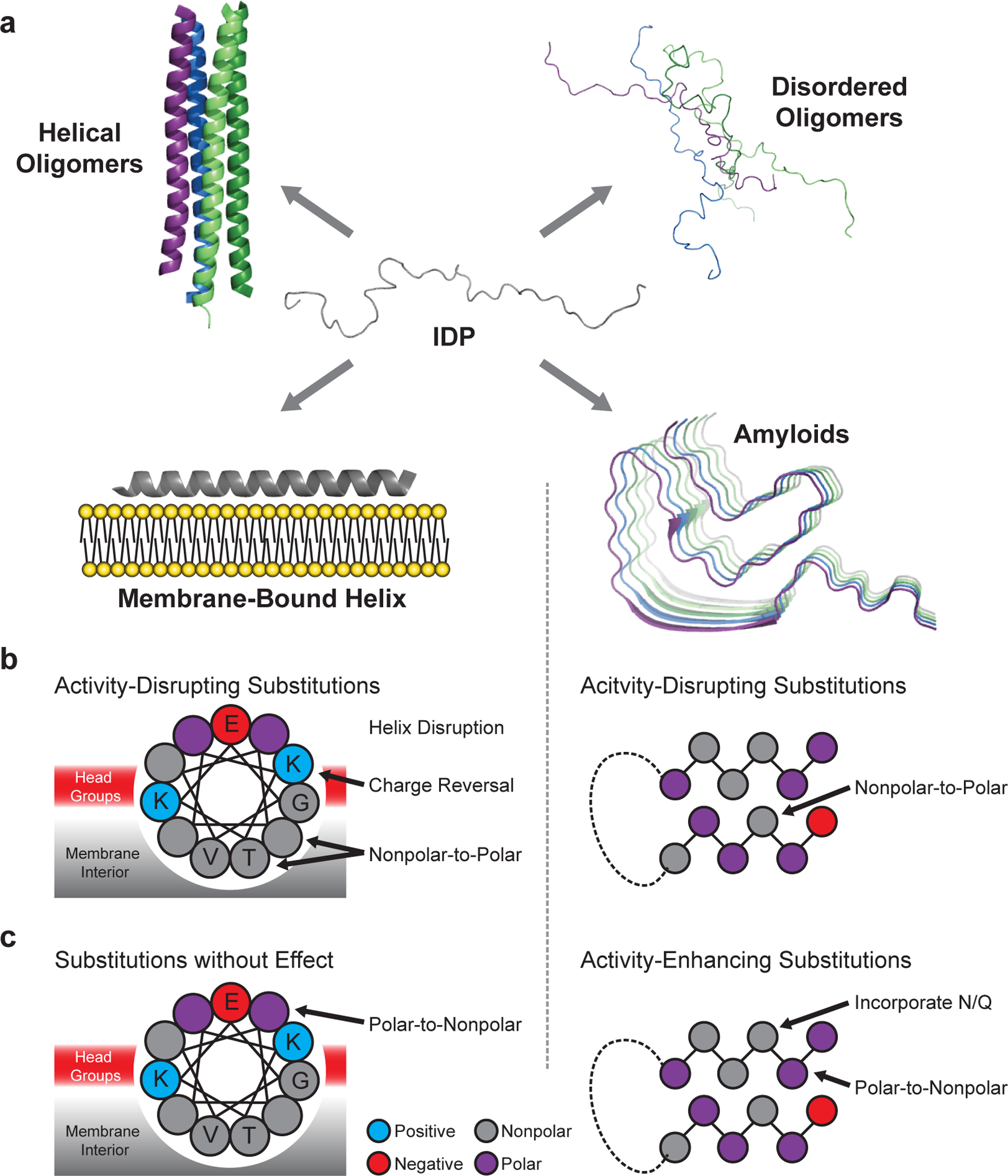
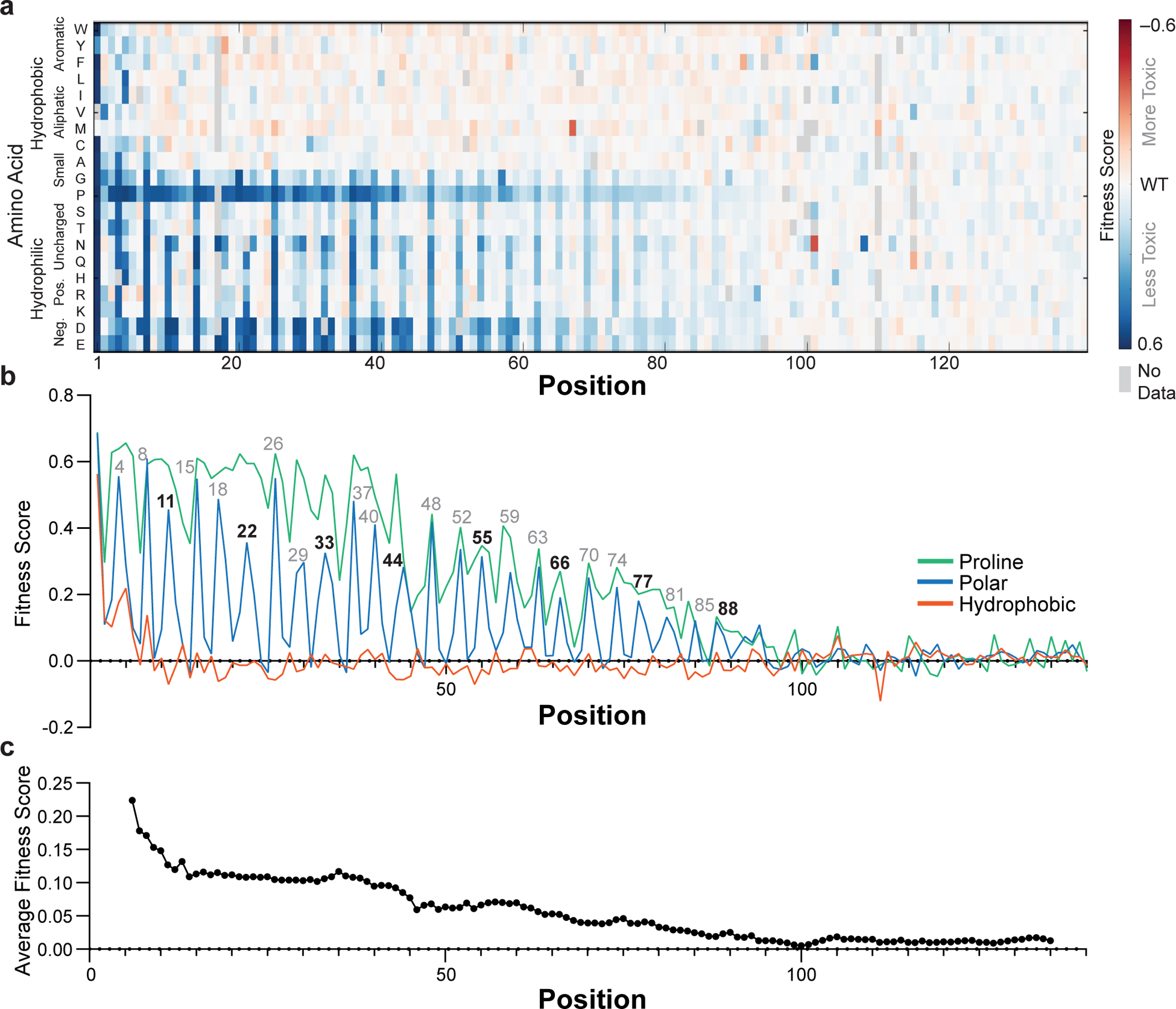
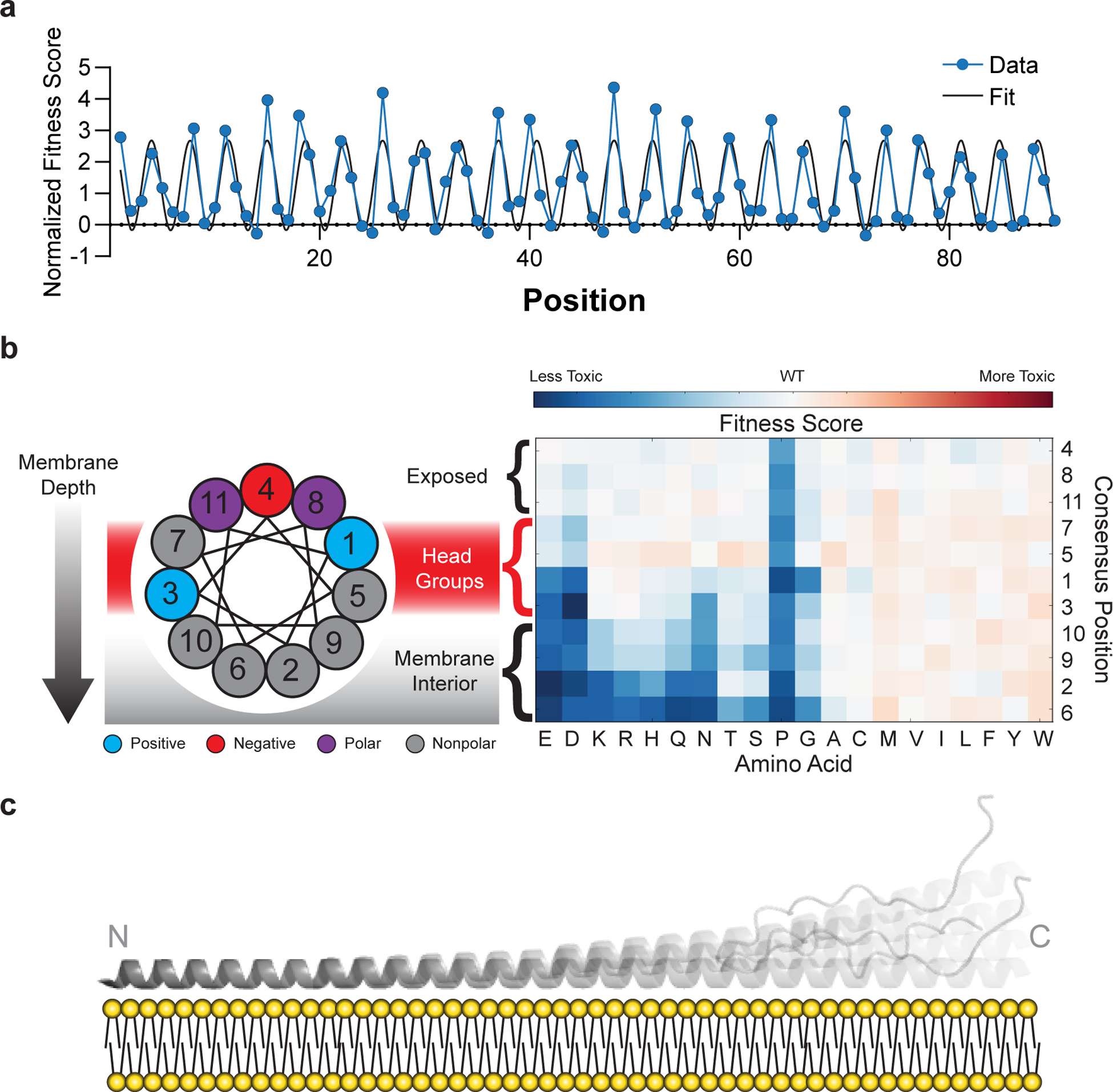
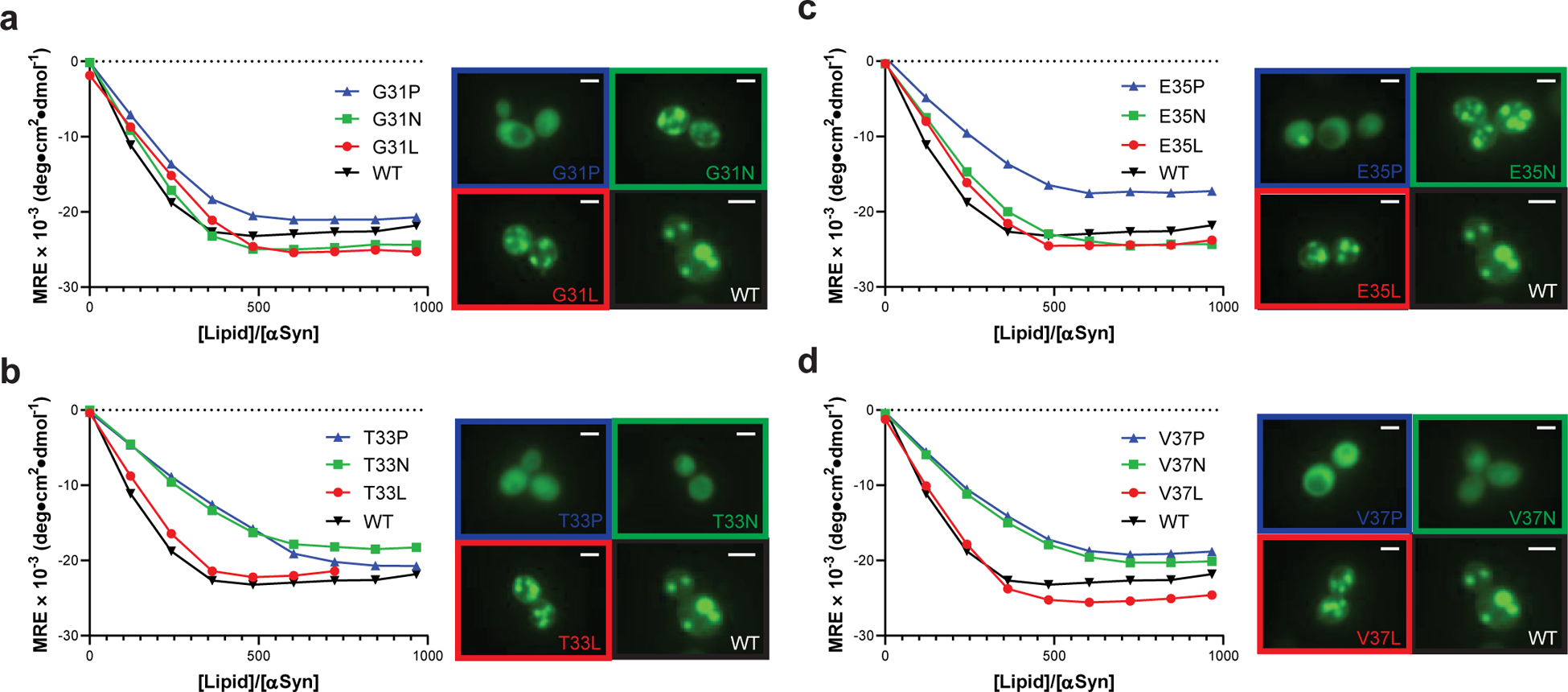
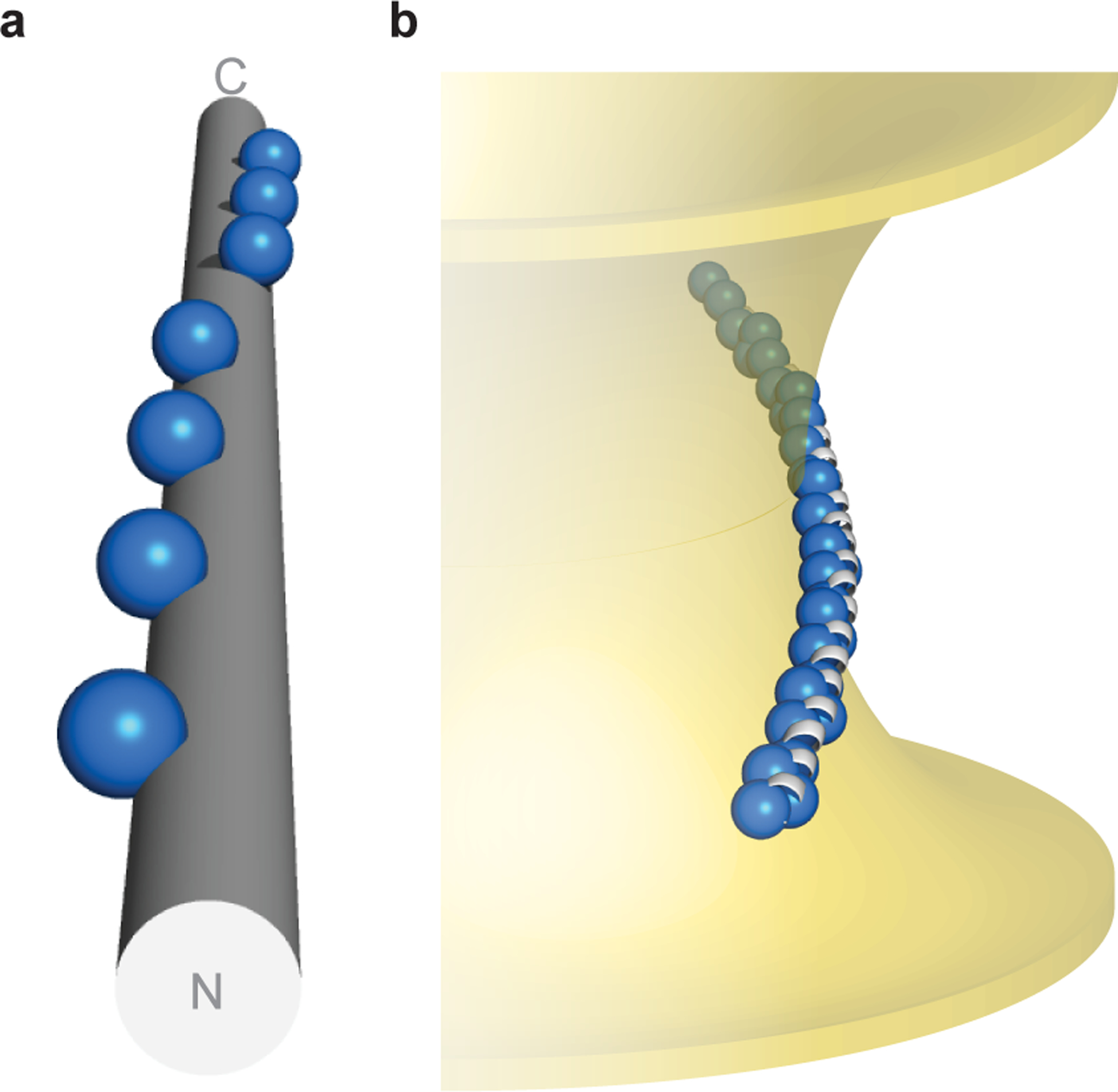
Defining the biologically active structures of proteins in their cellular environments remains challenging for proteins with multiple conformations and functions, where only a minor conformer might be associated with a given function. Here, we use deep mutational scanning to probe the structure and dynamics of α-synuclein, a protein known to adopt disordered, helical and amyloid conformations. We examined the effects of 2,600 single-residue substitutions on the ability of intracellularly expressed α-synuclein to slow the growth of yeast. Computational analysis of the data showed that the conformation responsible for this phenotype is a long, uninterrupted, amphiphilic helix with increasing dynamics toward the C terminus. Deep mutational scanning can therefore determine biologically active conformations in cellular environments, even for a highly dynamic multi-conformational protein.
Conflict of interest statement
Competing Interests
The authors declare no competing interests.
Figures
References
-
- Socolich M et al. Evolutionary information for specifying a protein fold. Nature 437, 512–518 (2005). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
