

Core components of DNA lagging strand synthesis machinery are essential for hepatitis B virus cccDNA formation
- PMID: 32152586
- PMCID: PMC7190442
- DOI: 10.1038/s41564-020-0678-0
Core components of DNA lagging strand synthesis machinery are essential for hepatitis B virus cccDNA formation
Abstract
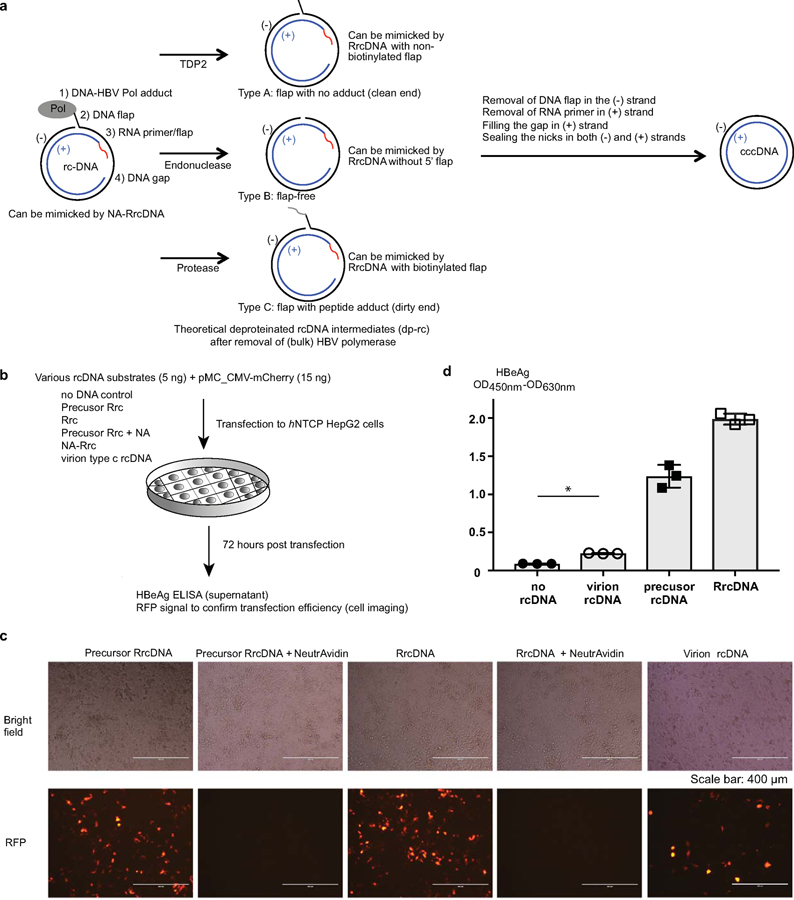
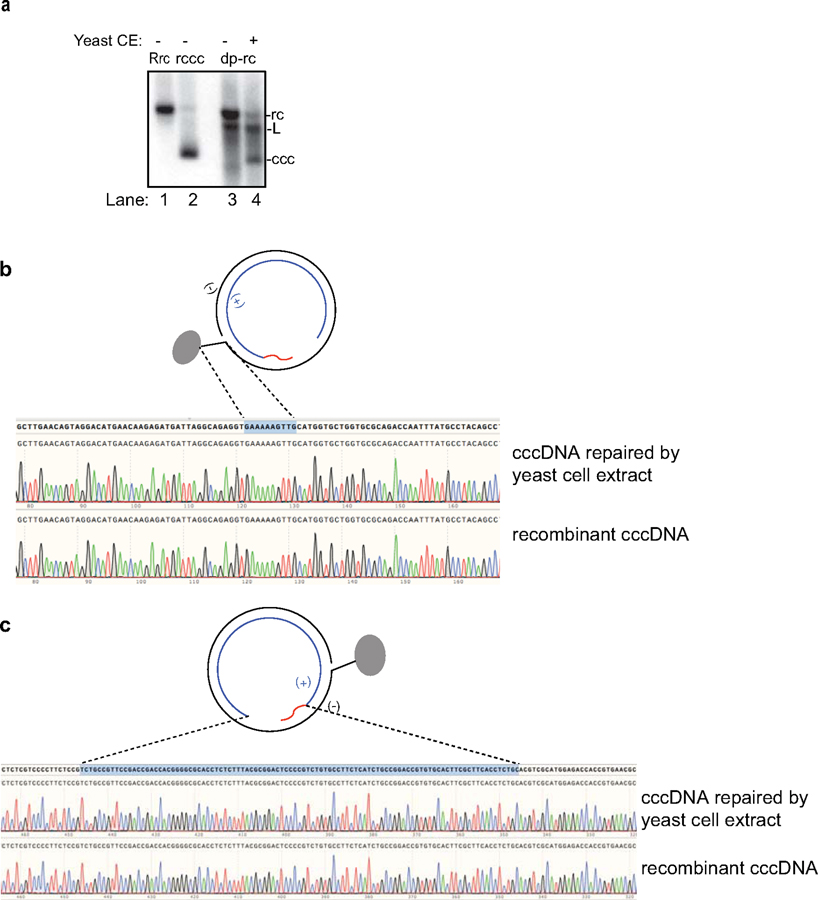
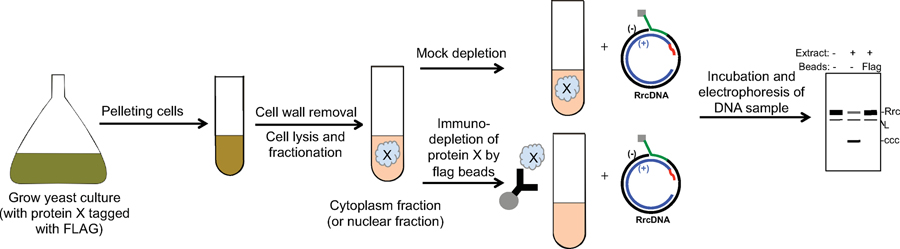
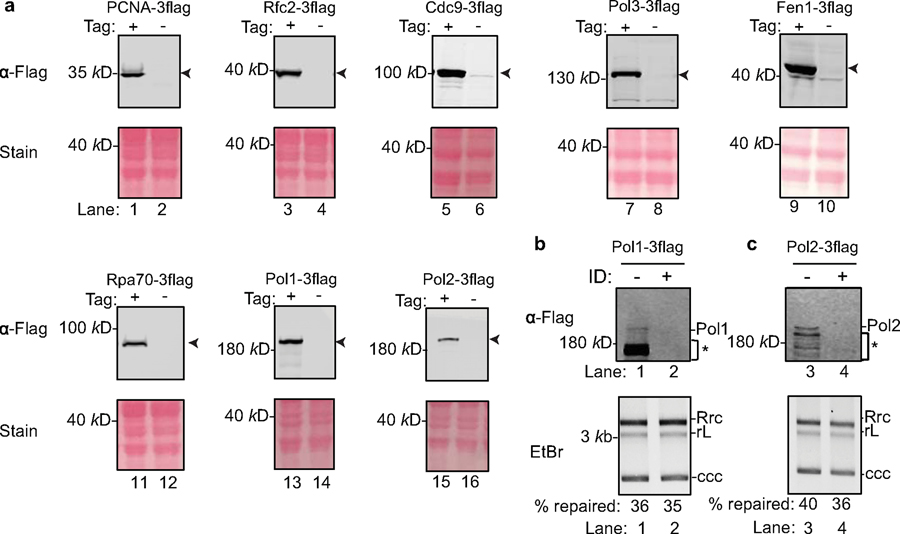
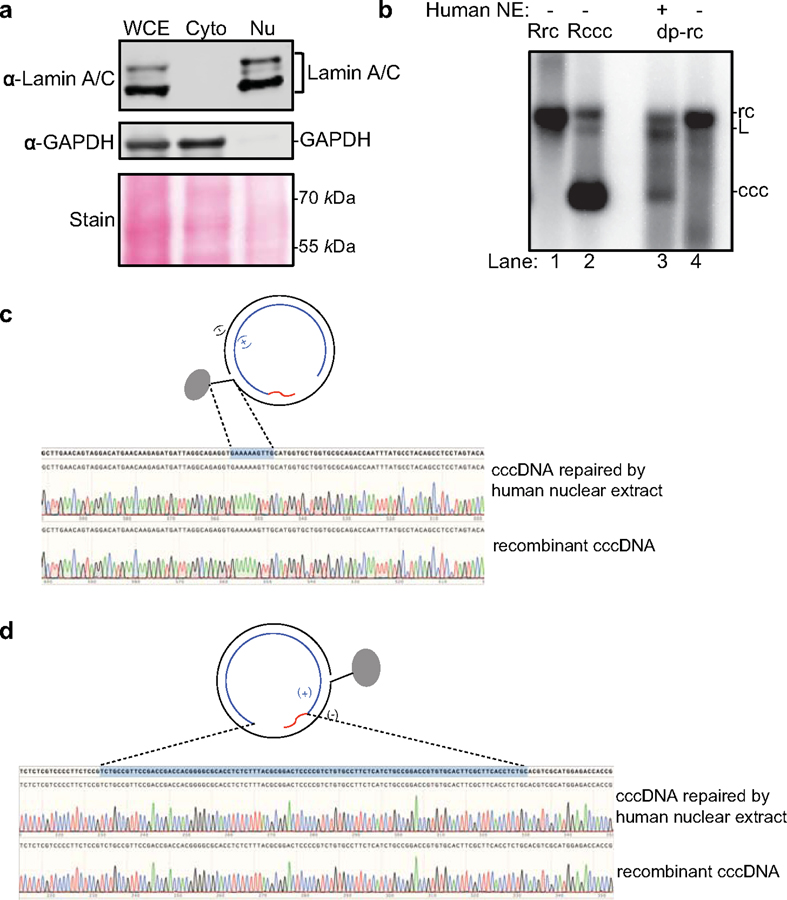
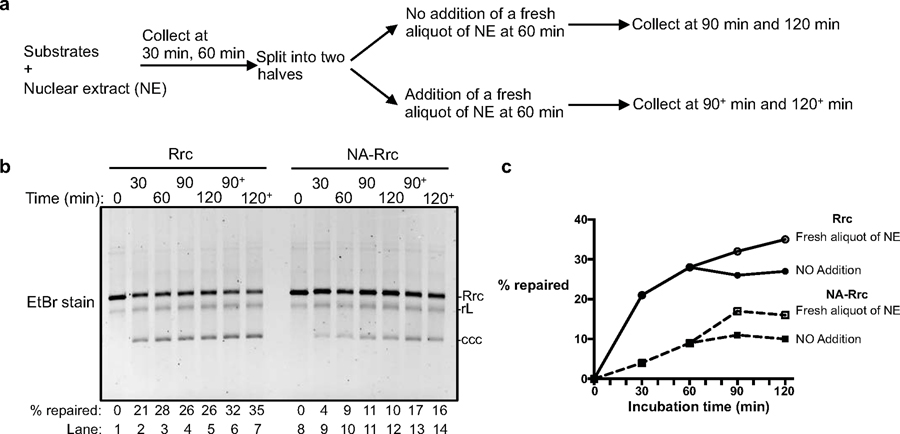
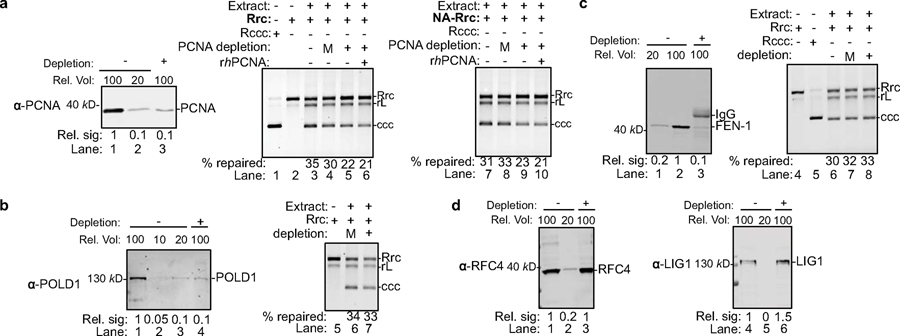
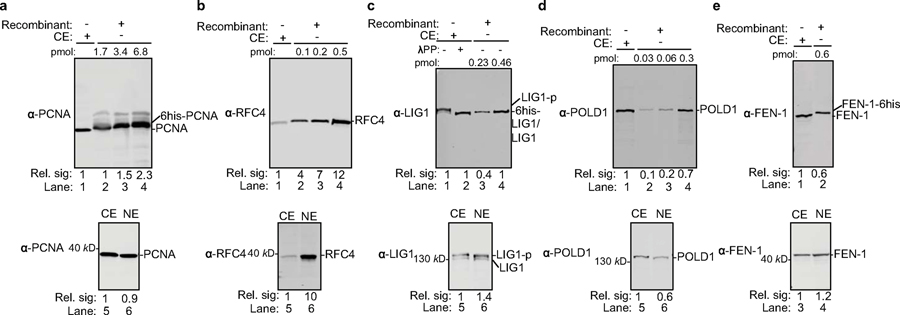
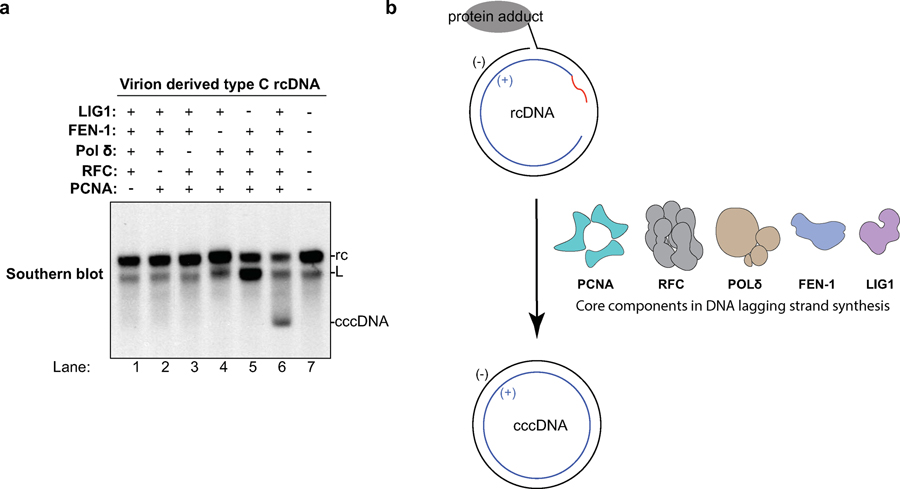
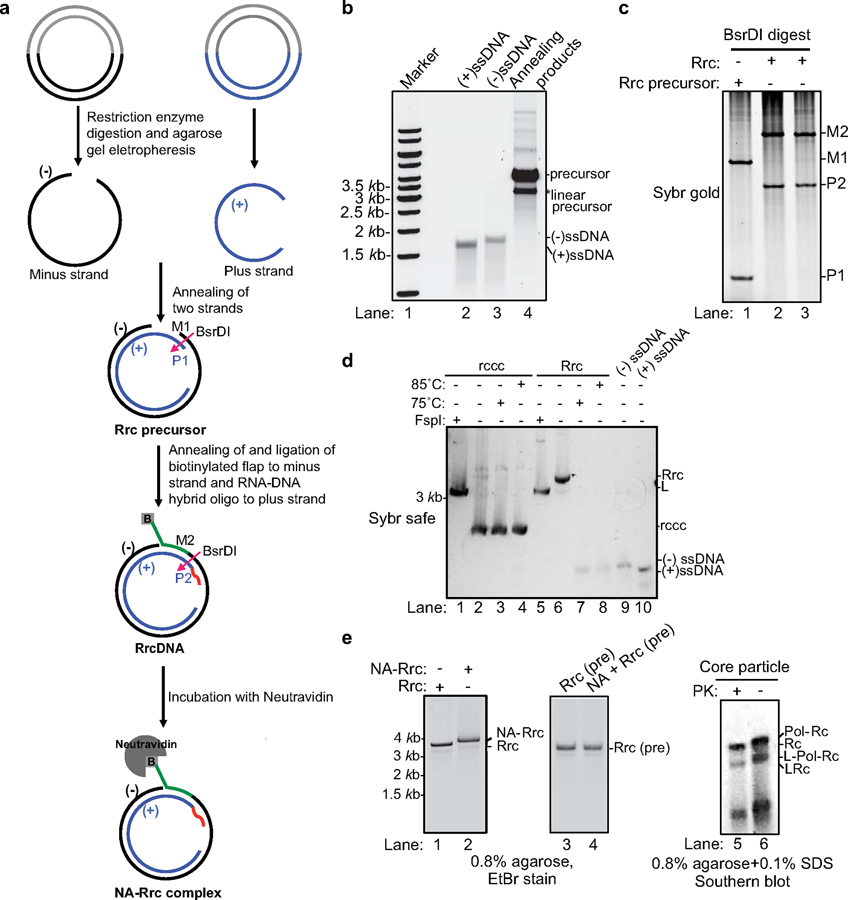
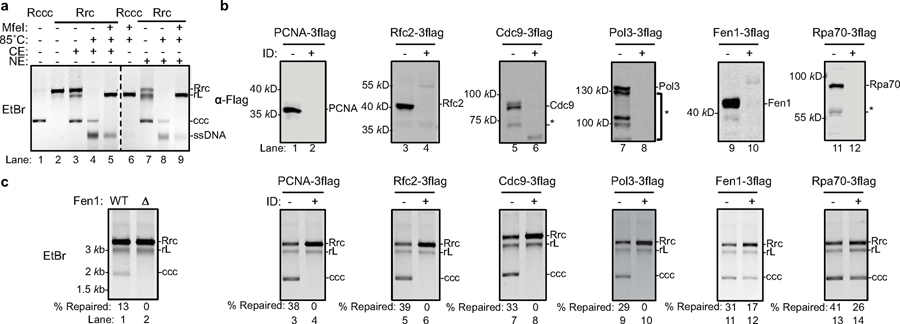
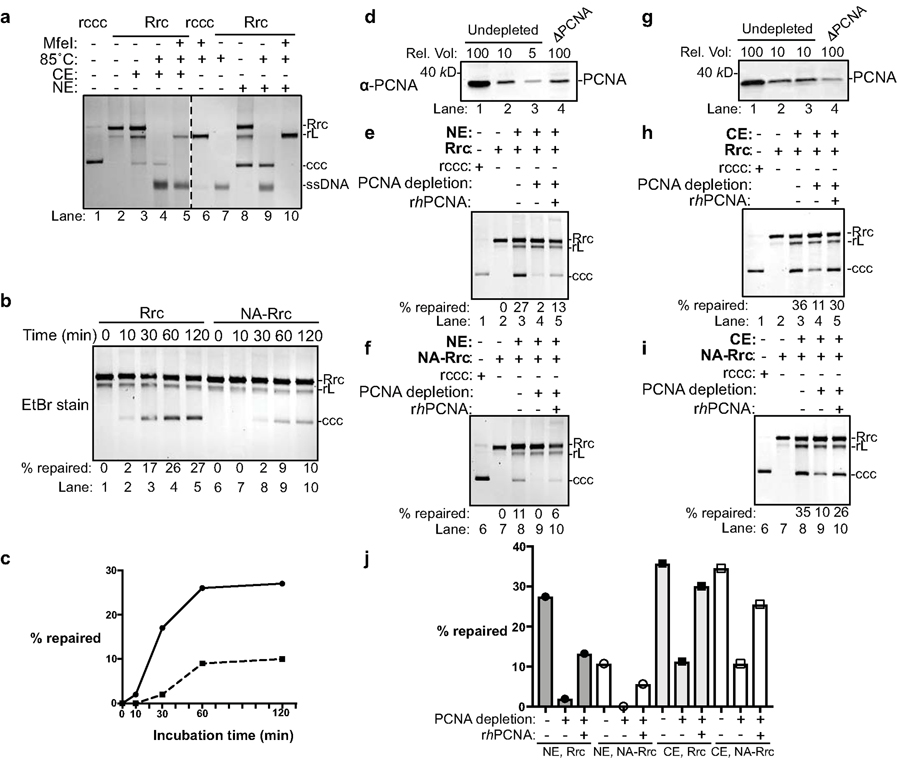
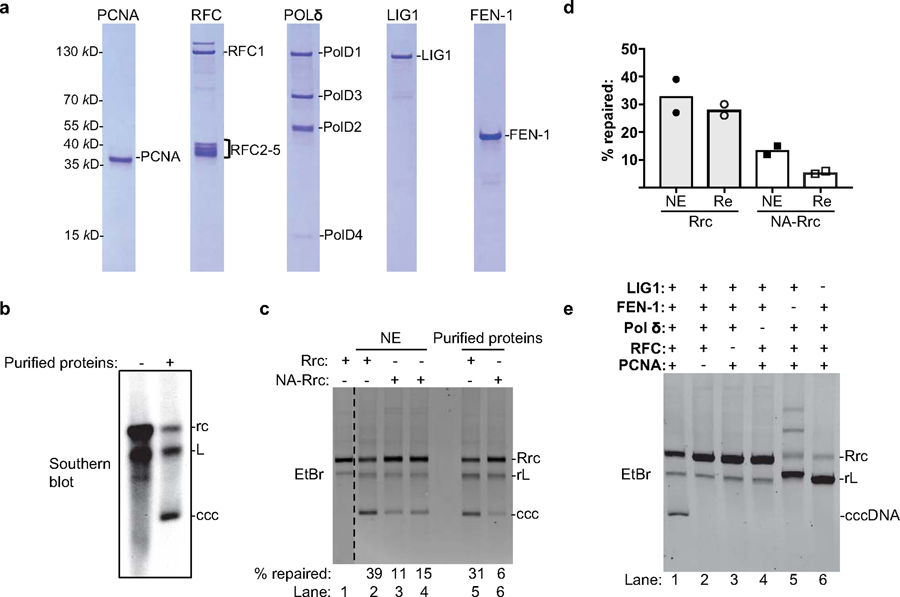
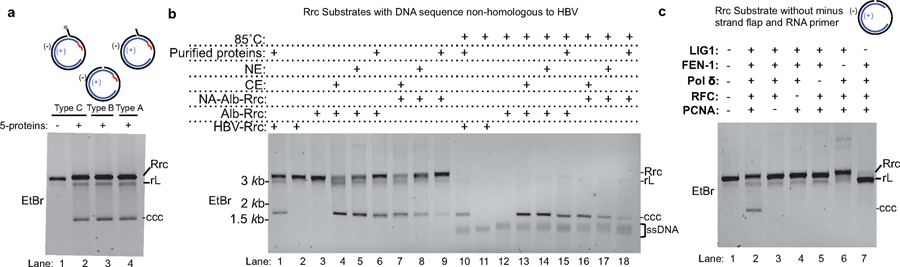
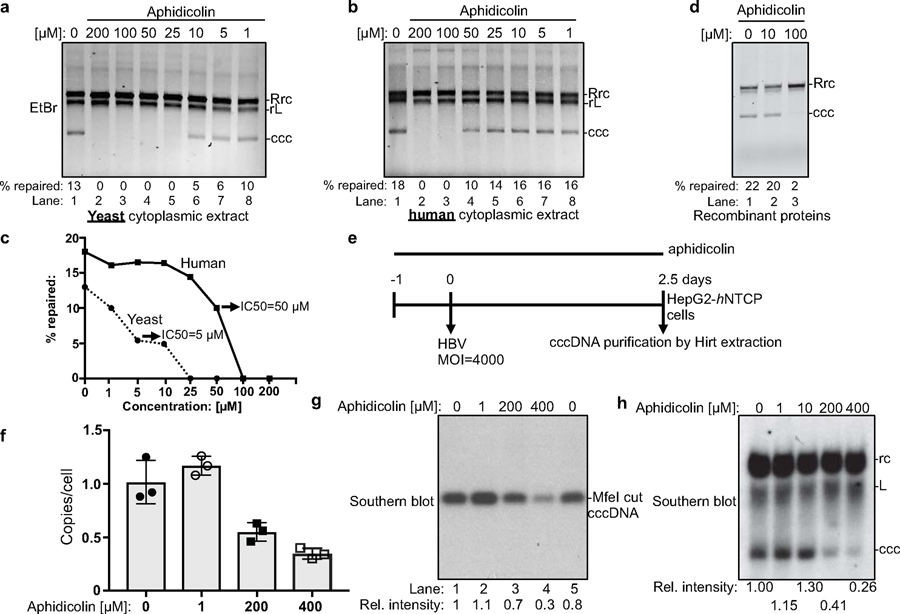
Chronic hepatitis B virus (HBV) infections result in 887,000 deaths annually. The central challenge in curing HBV is eradication of the stable covalently closed circular DNA (cccDNA) form of the viral genome, which is formed by the repair of lesion-bearing HBV relaxed circular DNA delivered by the virions to hepatocytes. The complete and minimal set of host factors involved in cccDNA formation is unknown, largely due to the lack of a biochemical system that fully reconstitutes cccDNA formation. Here, we have developed experimental systems where various HBV relaxed-circular-DNA substrates are repaired to form cccDNA by both cell extracts and purified human proteins. Using yeast- and human-extract screenings, we identified five core components of lagging-strand synthesis as essential for cccDNA formation: proliferating cell nuclear antigen, the replication factor C complex, DNA polymerase δ, flap endonuclease 1 and DNA ligase 1. We reconstituted cccDNA formation with purified human homologues, establishing these as a minimal set of factors for cccDNA formation. We further demonstrated that treatment with the DNA-polymerase inhibitor aphidicolin diminishes cccDNA formation both in biochemical assays and in HBV-infected human cells. Together, our findings define key components in HBV cccDNA formation.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
Comment in
-
Have the Starting Lineup of Five for Hepatitis B Virus Covalently Closed Circular DNA Synthesis Been Identified?Hepatology. 2020 Sep;72(3):1142-1144. doi: 10.1002/hep.31408. Hepatology. 2020. PMID: 32502295 No abstract available.
References
-
- Nassal M. HBV cccDNA: viral persistence reservoir and key obstacle for a cure of chronic hepatitis B. Gut 64, 1972–1984 (2015). - PubMed
-
- Wang GH & Seeger C. The reverse transcriptase of hepatitis B virus acts as a protein primer for viral DNA synthesis. Cell 71, 663–670 (1992). - PubMed
-
- Gerlich WH & Robinson WS. Hepatitis B virus contains protein attached to the 5’ terminus of its complete DNA strand. Cell 21, 801–809 (1980). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
