

NELF Regulates a Promoter-Proximal Step Distinct from RNA Pol II Pause-Release
- PMID: 32155413
- PMCID: PMC7402197
- DOI: 10.1016/j.molcel.2020.02.014
NELF Regulates a Promoter-Proximal Step Distinct from RNA Pol II Pause-Release
Abstract
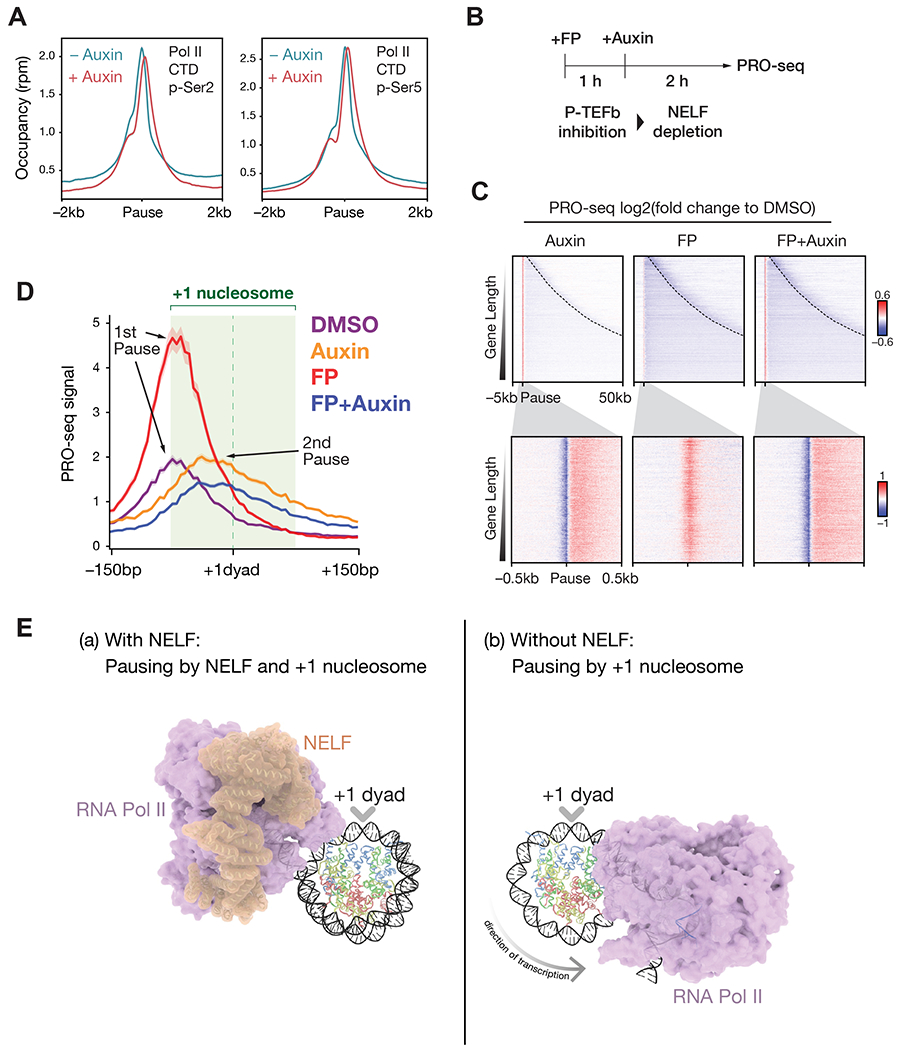
RNA polymerase II (RNA Pol II) is generally paused at promoter-proximal regions in most metazoans, and based on in vitro studies, this function has been attributed to the negative elongation factor (NELF). Here, we show that upon rapid depletion of NELF, RNA Pol II fails to be released into gene bodies, stopping instead around the +1 nucleosomal dyad-associated region. The transition to the 2nd pause region is independent of positive transcription elongation factor P-TEFb. During the heat shock response, RNA Pol II is rapidly released from pausing at heat shock-induced genes, while most genes are paused and transcriptionally downregulated. Both of these aspects of the heat shock response remain intact upon NELF loss. We find that NELF depletion results in global loss of cap-binding complex from chromatin without global reduction of nascent transcript 5' cap stability. Thus, our studies implicate NELF functioning in early elongation complexes distinct from RNA Pol II pause-release.
Keywords: NELF; PRO-cap; PRO-seq; RNA Polymerase II; cap-binding complex; m7G cap; mRNA capping; promoter-proximal pausing; super elongation complex; transcription elongation.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures






References
-
- Aiyar SE, Sun J-L, Blair AL, Moskaluk CA, Lu Y-Z, Ye Q-N, Yamaguchi Y, Mukherjee A, Ren D-M, Handa H, et al. (2004). Attenuation of estrogen receptor alpha-mediated transcription through estrogen-stimulated recruitment of a negative elongation factor. Genes & Development 18, 2134–2146. - PMC - PubMed
-
- Aprile-Garcia F, Tomar P, Hummel B, Khavaran A, and Sawarkar R (2019). Nascent-protein ubiquitination is required for heat shock–induced gene downregulation in human cells. Nat. Struct. Mol. Biol 26, 1–16. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
