

Specific Recruitment of Phosphoinositide Species to the Plant-Pathogen Interfacial Membrane Underlies Arabidopsis Susceptibility to Fungal Infection
- PMID: 32156686
- PMCID: PMC7203932
- DOI: 10.1105/tpc.19.00970
Specific Recruitment of Phosphoinositide Species to the Plant-Pathogen Interfacial Membrane Underlies Arabidopsis Susceptibility to Fungal Infection
Abstract
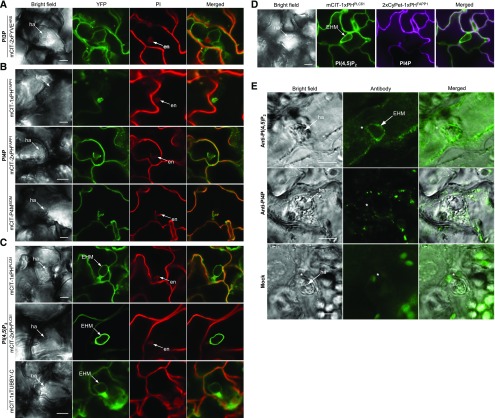
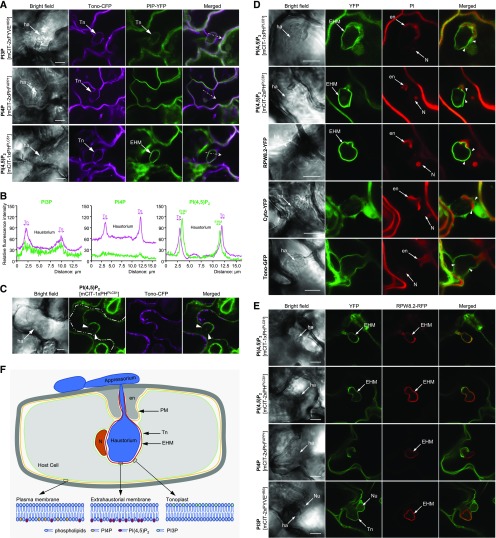
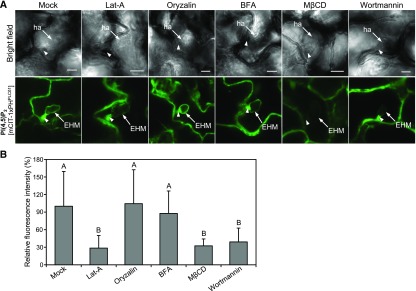
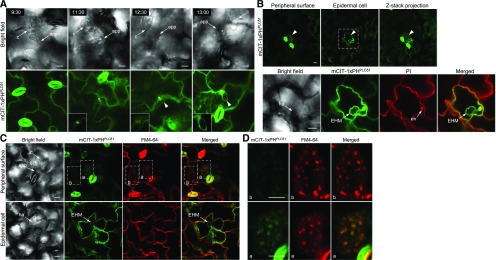
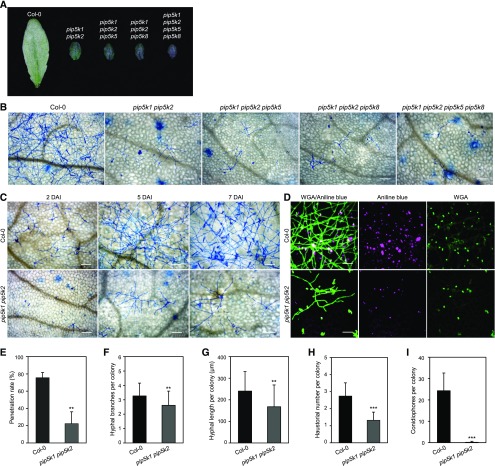
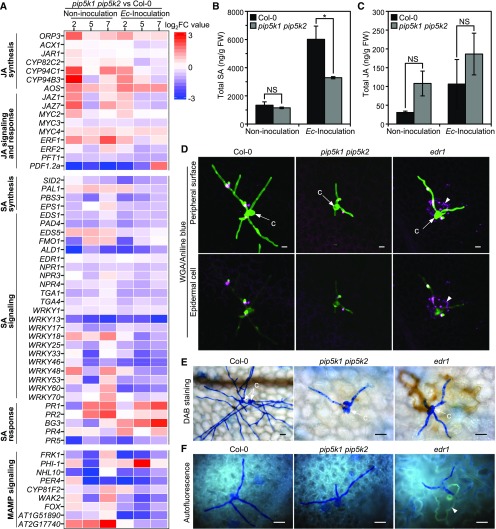
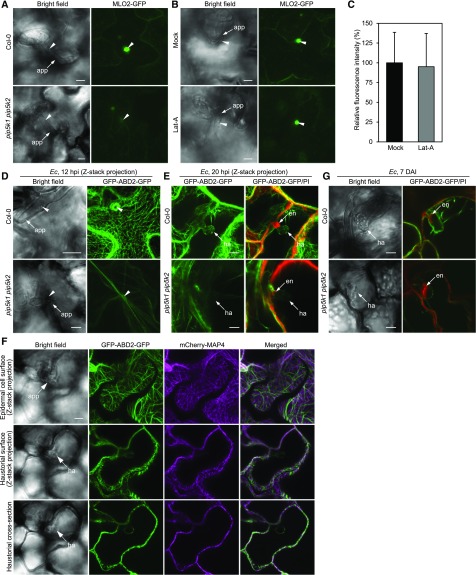
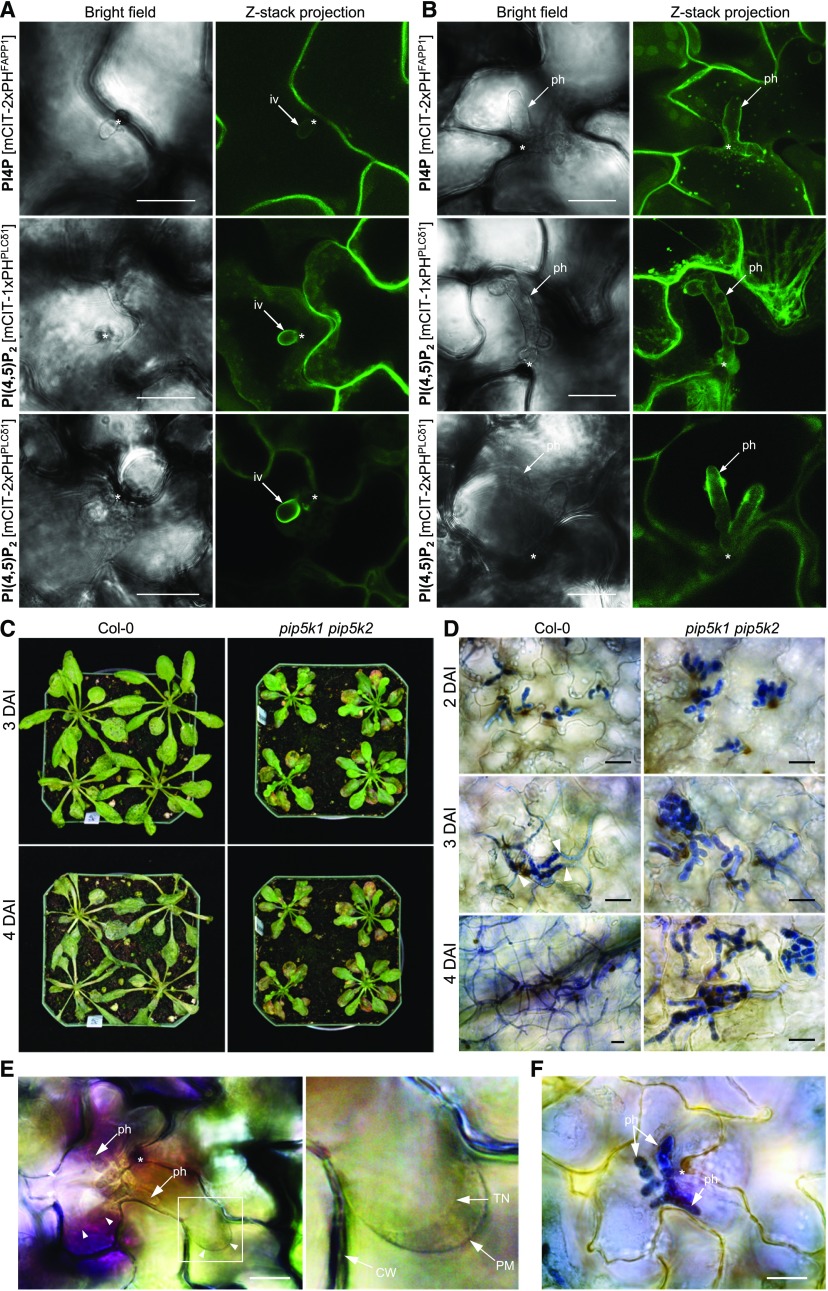
Different phosphoinositides enriched at the membranes of specific subcellular compartments within plant cells contribute to organelle identity, ensuring appropriate cellular trafficking and function. During the infection of plant cells, biotrophic pathogens such as powdery mildews enter plant cells and differentiate into haustoria. Each haustorium is enveloped by an extrahaustorial membrane (EHM) derived from the host plasma membrane. Little is known about the EHM biogenesis and identity. Here, we demonstrate that among the two plasma membrane phosphoinositides in Arabidopsis (Arabidopsis thaliana), PI(4,5)P2 is dynamically up-regulated at powdery mildew infection sites and recruited to the EHM, whereas PI4P is absent in the EHM. Lateral transport of PI(4,5)P2 into the EHM occurs through a brefeldin A-insensitive but actin-dependent trafficking pathway. Furthermore, the lower levels of PI(4,5)P2 in pip5k1 pip5k2 mutants inhibit fungal pathogen development and cause disease resistance, independent of cell death-associated defenses and involving impaired host susceptibility. Our results reveal that plant biotrophic and hemibiotrophic pathogens modulate the subcellular distribution of host phosphoinositides and recruit PI(4,5)P2 as a susceptibility factor for plant disease.
© 2020 American Society of Plant Biologists. All rights reserved.
Figures

Comment in
-
Good Fats, Bad Fats: Phosphoinositide Species Differentially Localize to Plant-Pathogen Interfaces and Influence Disease Progression.Plant Cell. 2020 May;32(5):1355-1356. doi: 10.1105/tpc.20.00193. Epub 2020 Mar 13. Plant Cell. 2020. PMID: 32169956 Free PMC article. No abstract available.
References
-
- Asai T., Tena G., Plotnikova J., Willmann M.R., Chiu W.-L., Gomez-Gomez L., Boller T., Ausubel F.M., Sheen J.(2002). MAP kinase signalling cascade in Arabidopsis innate immunity. Nature 415: 977–983. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
