

TAp63-Regulated miRNAs Suppress Cutaneous Squamous Cell Carcinoma through Inhibition of a Network of Cell-Cycle Genes
- PMID: 32156775
- PMCID: PMC7299759
- DOI: 10.1158/0008-5472.CAN-19-1892
TAp63-Regulated miRNAs Suppress Cutaneous Squamous Cell Carcinoma through Inhibition of a Network of Cell-Cycle Genes
Abstract
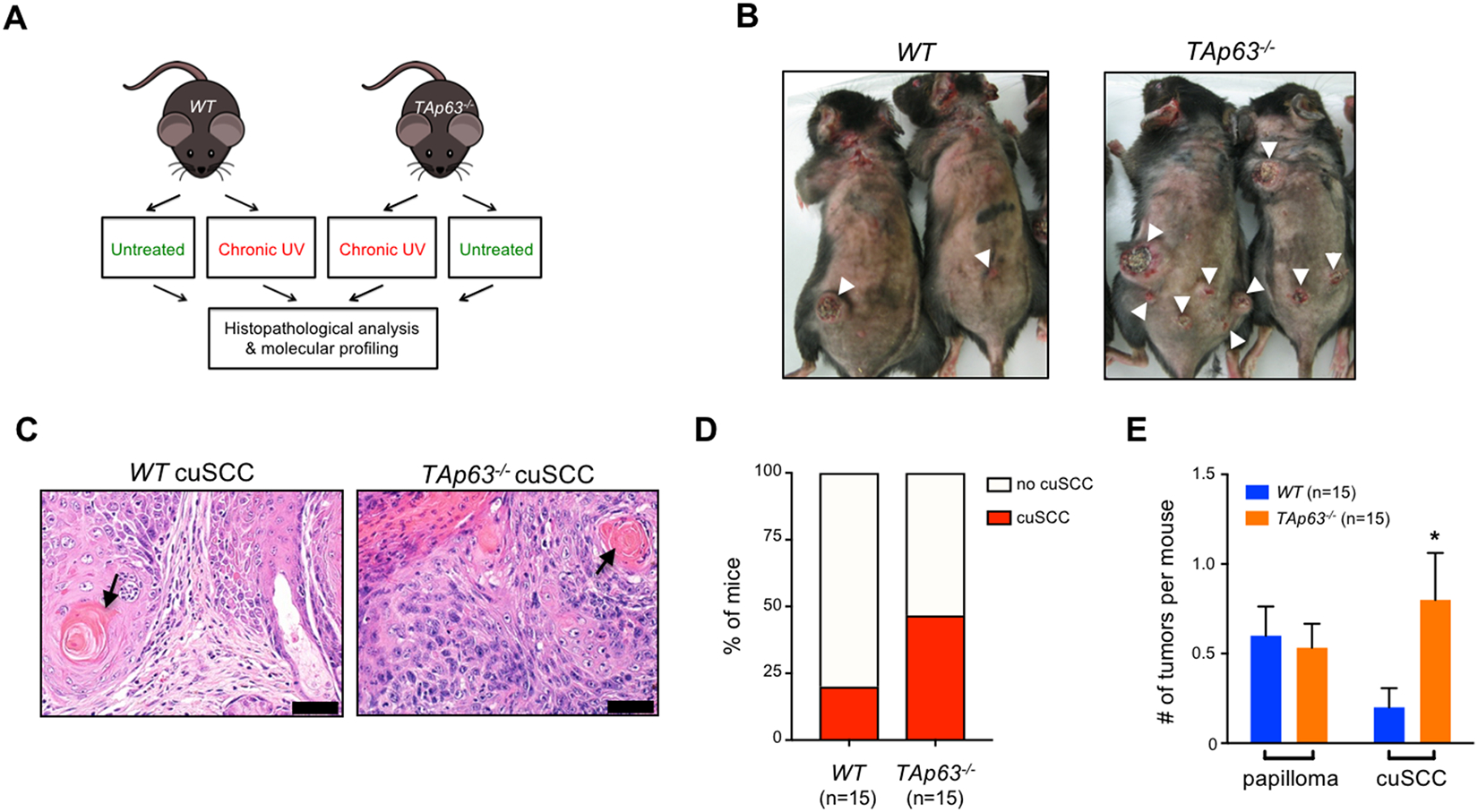
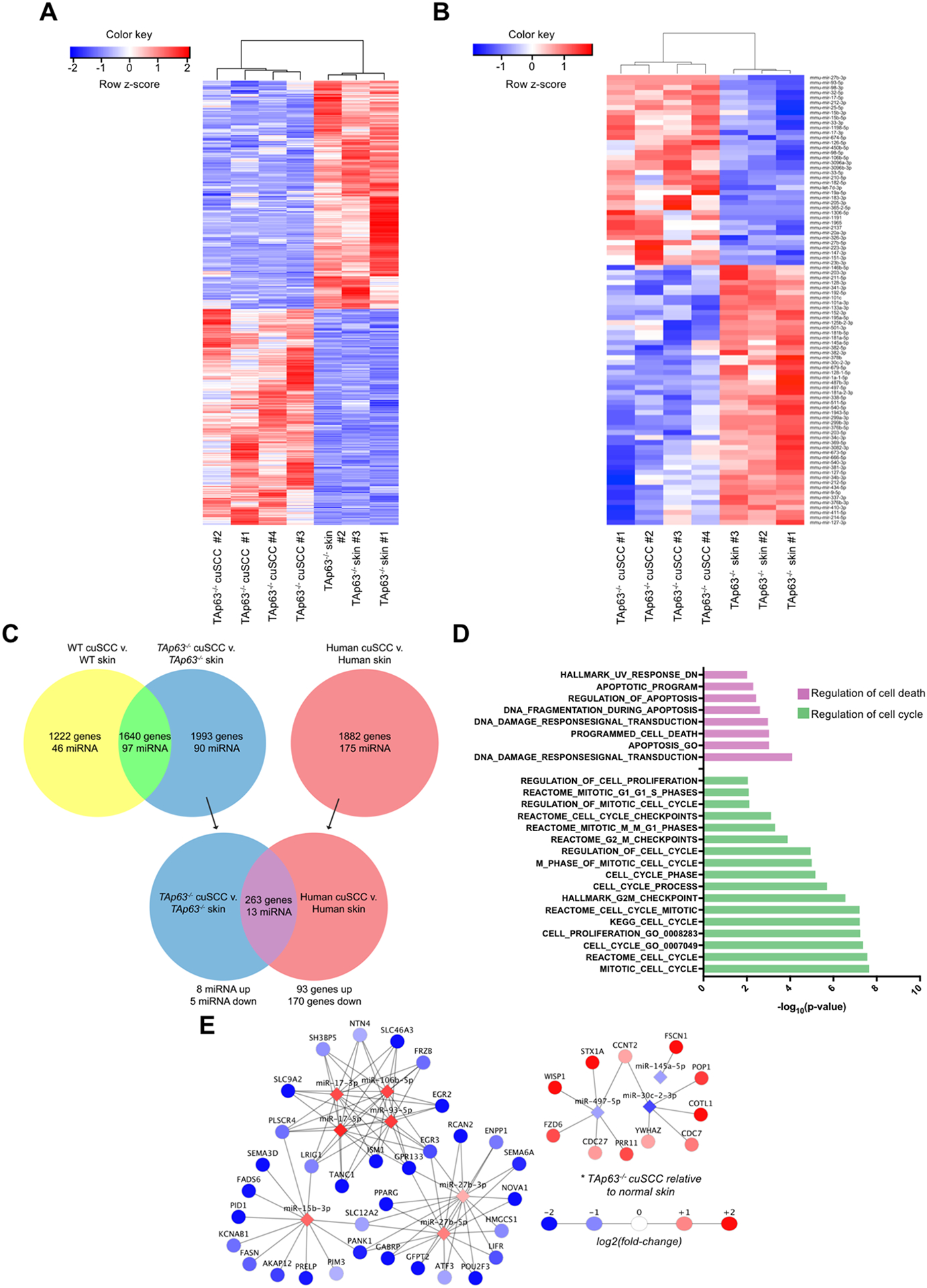
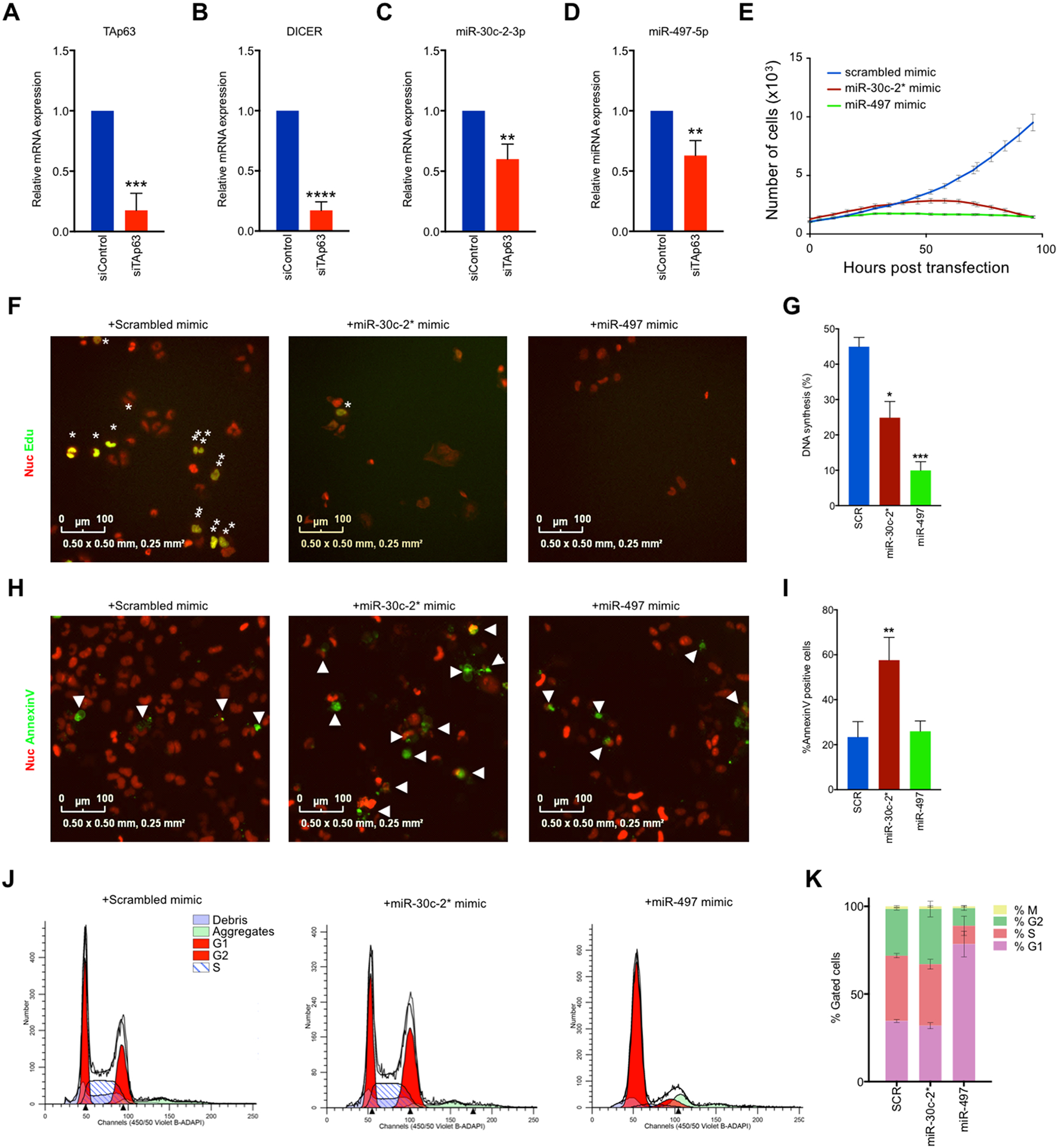
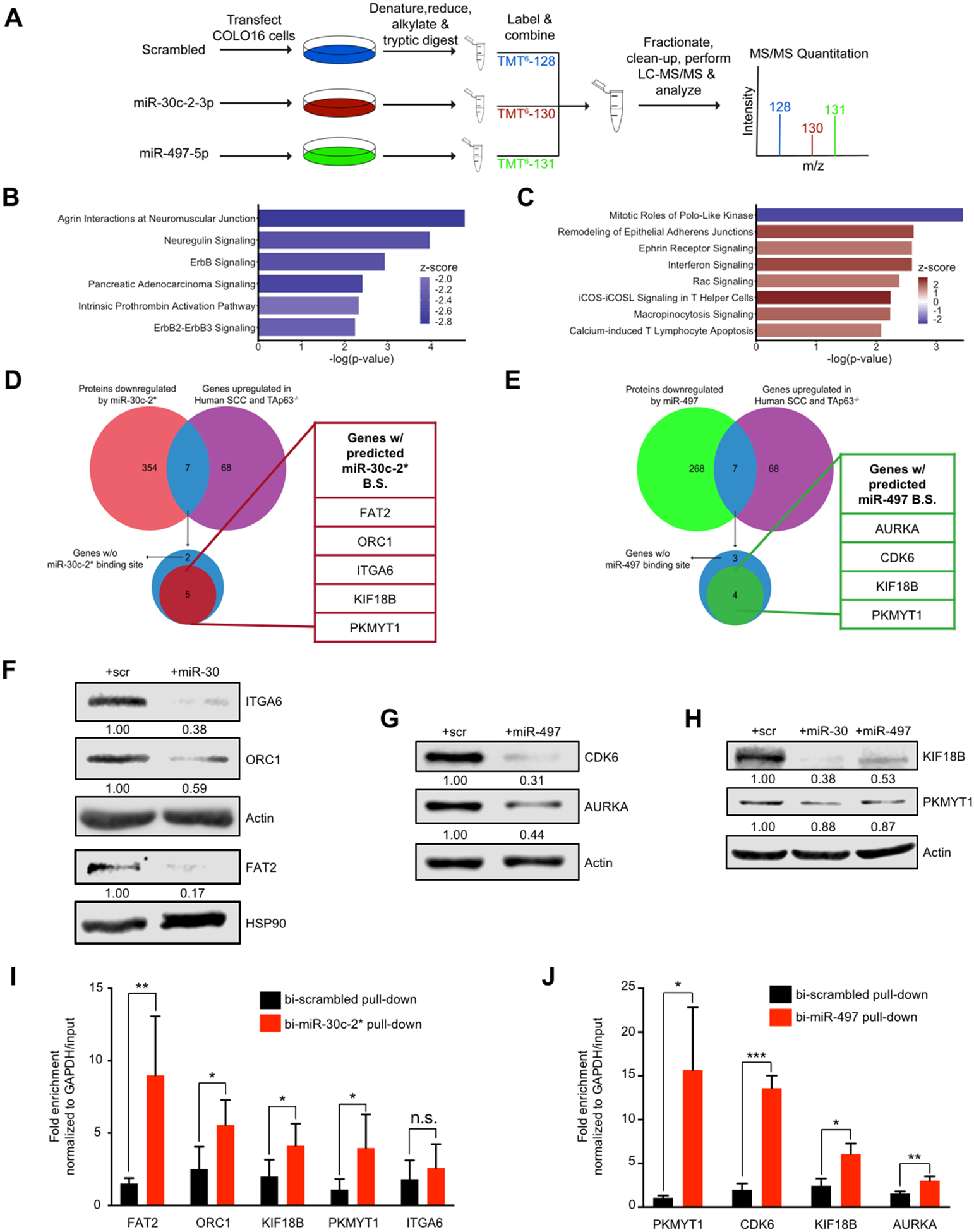
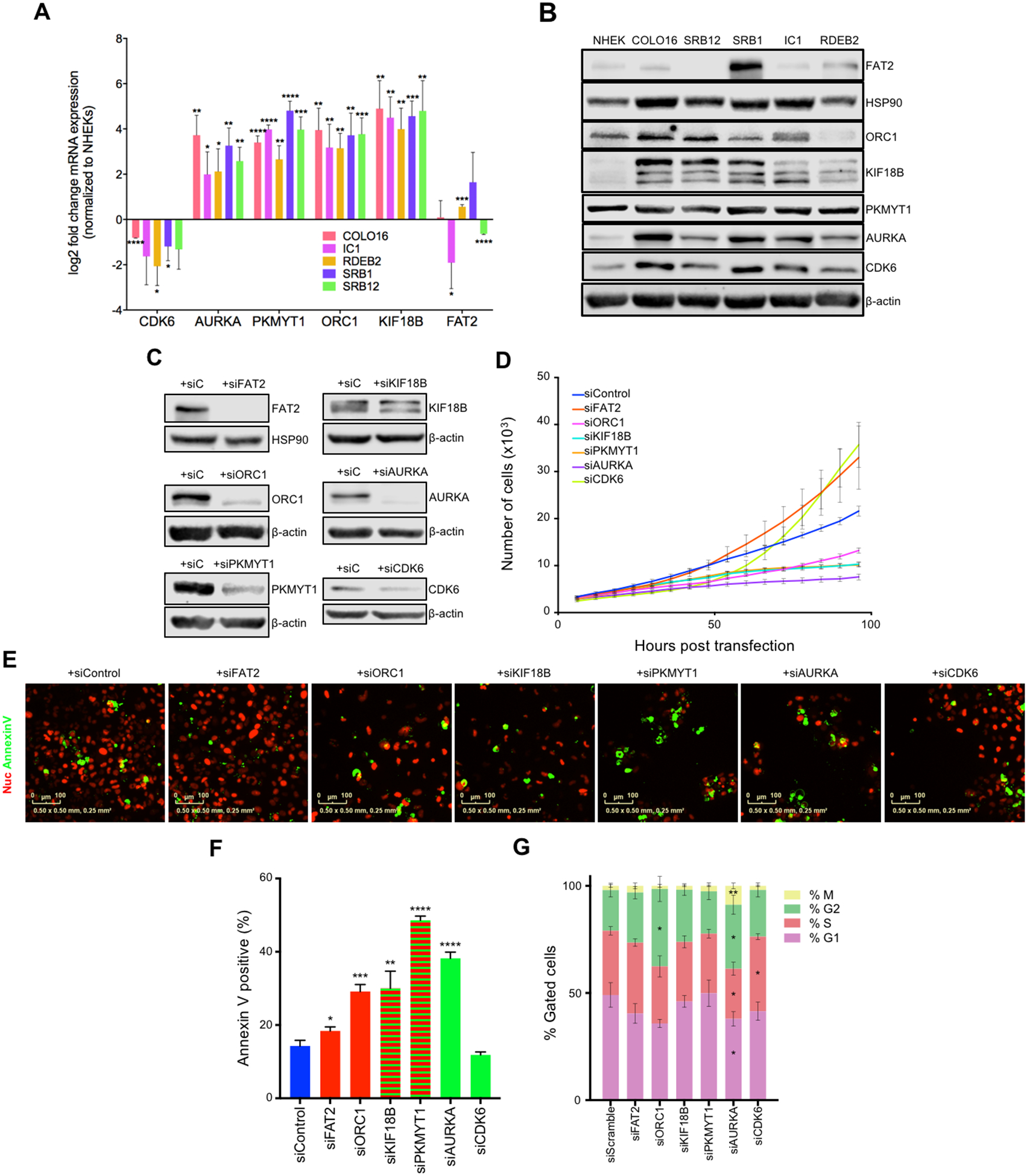
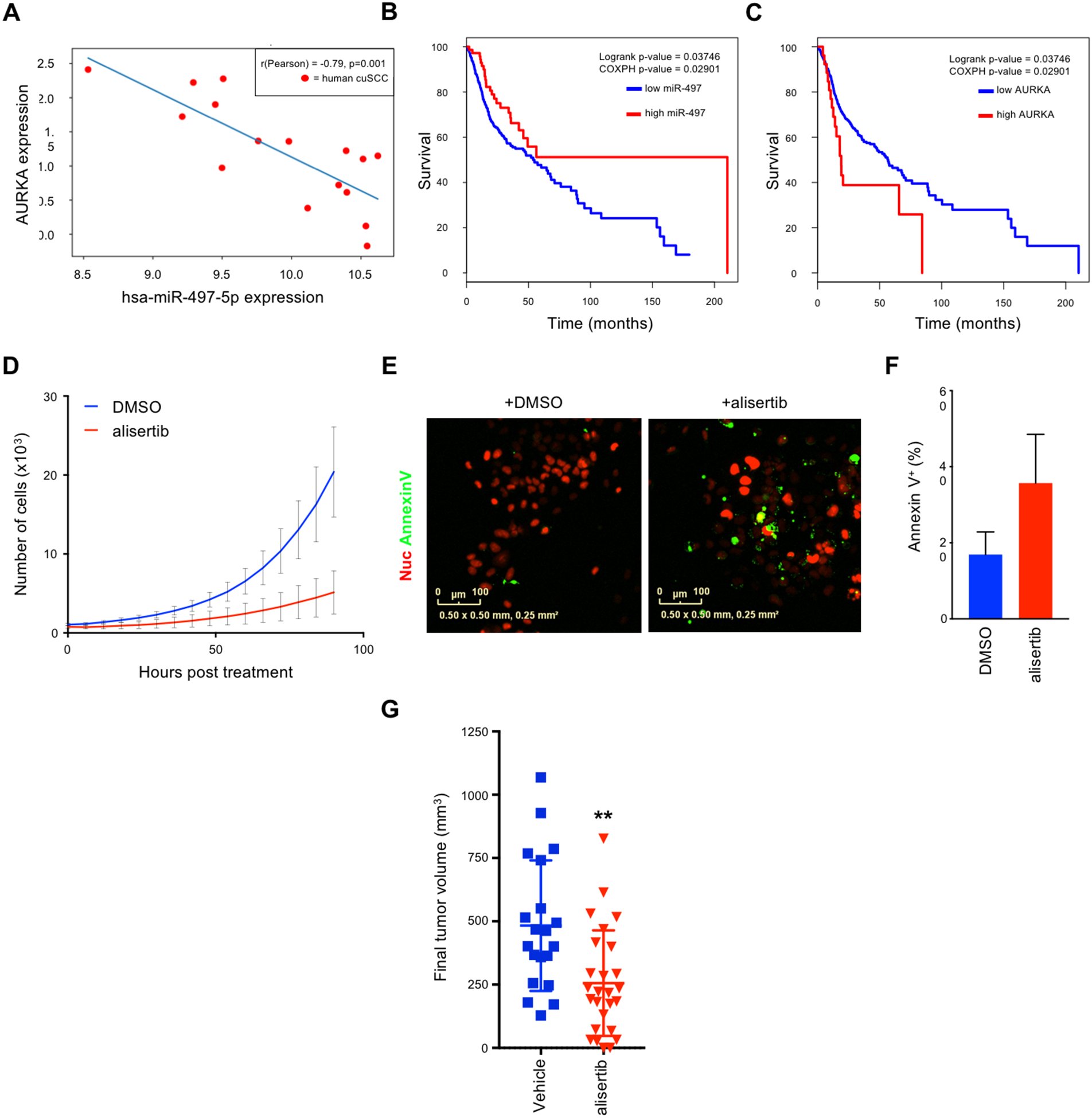
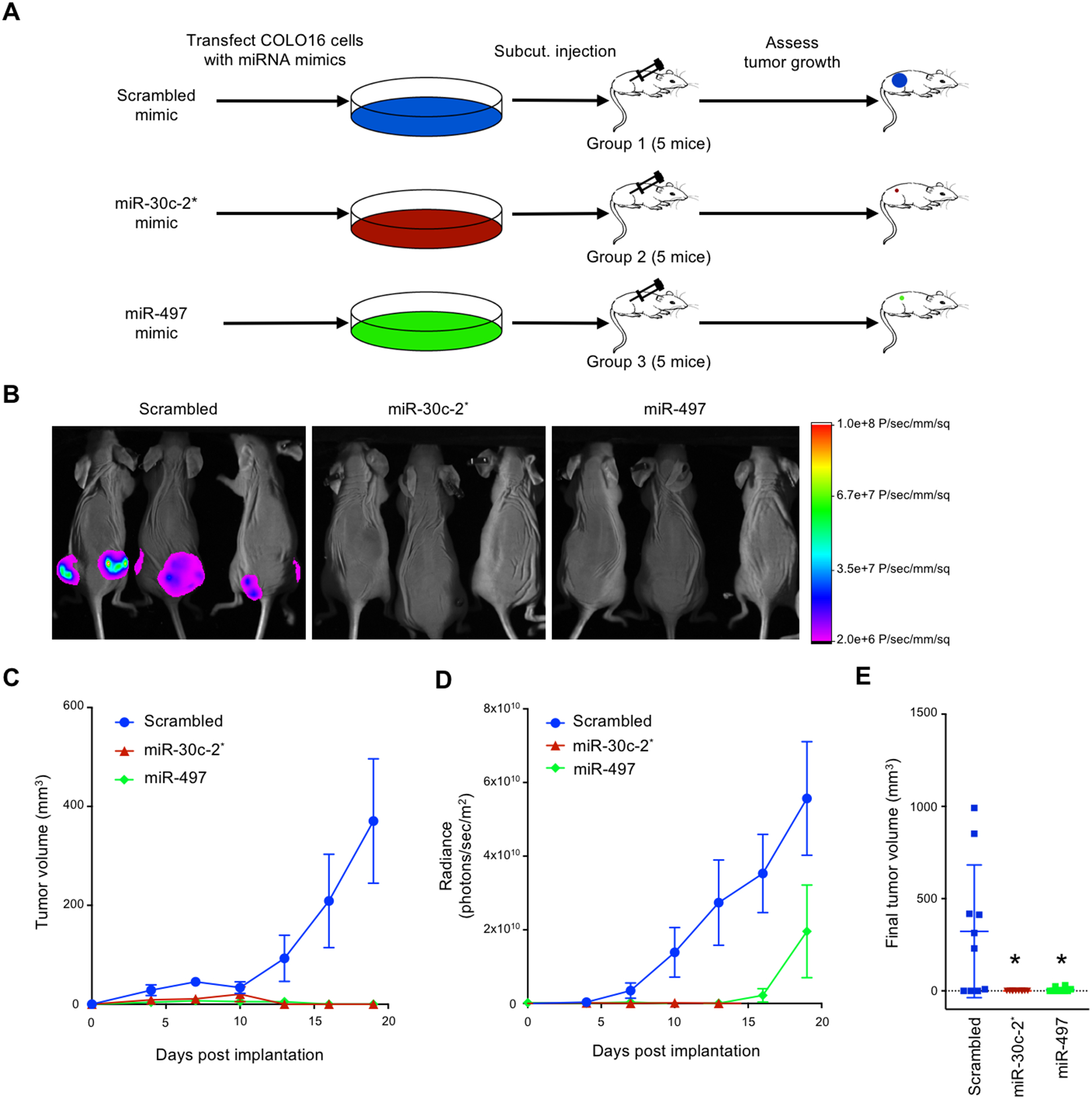
TAp63 is a p53 family member and potent tumor and metastasis suppressor. Here, we show that TAp63-/- mice exhibit an increased susceptibility to ultraviolet radiation-induced cutaneous squamous cell carcinoma (cuSCC). A human-to-mouse comparison of cuSCC tumors identified miR-30c-2* and miR-497 as underexpressed in TAp63-deficient cuSCC. Reintroduction of these miRNAs significantly inhibited the growth of cuSCC cell lines and tumors. Proteomic profiling of cells expressing either miRNA showed downregulation of cell-cycle progression and mitosis-associated proteins. A mouse to human and cross-platform comparison of RNA-sequencing and proteomics data identified a 7-gene signature, including AURKA, KIF18B, PKMYT1, and ORC1, which were overexpressed in cuSCC. Knockdown of these factors in cuSCC cell lines suppressed tumor cell proliferation and induced apoptosis. In addition, selective inhibition of AURKA suppressed cuSCC cell proliferation, induced apoptosis, and showed antitumor effects in vivo. Finally, treatment with miR-30c-2* or miR-497 miRNA mimics was highly effective in suppressing cuSCC growth in vivo. Our data establish TAp63 as an essential regulator of novel miRNAs that can be therapeutically targeted for potent suppression of cuSCC. SIGNIFICANCE: This study provides preclinical evidence for the use of miR-30c-2*/miR-497 delivery and AURKA inhibition in the treatment of cuSCC, which currently has no FDA-approved targeted therapies.See related commentary by Parrales and Iwakuma, p. 2439.
©2020 American Association for Cancer Research.
Conflict of interest statement
Conflict of interest: The authors have declared that no conflict of interest exists.
Figures
Comment in
-
TAp63-miRNA-AURKA Axis as a Therapeutic Target for Cutaneous Squamous Cell Carcinoma.Cancer Res. 2020 Jun 15;80(12):2439-2440. doi: 10.1158/0008-5472.CAN-20-1215. Cancer Res. 2020. PMID: 32540854 Free PMC article.
Comment on
-
TAp63-miRNA-AURKA Axis as a Therapeutic Target for Cutaneous Squamous Cell Carcinoma.Cancer Res. 2020 Jun 15;80(12):2439-2440. doi: 10.1158/0008-5472.CAN-20-1215. Cancer Res. 2020. PMID: 32540854 Free PMC article.
References
-
- Rogers HW, Weinstock MA, Harris AR, Hinckley MR, Feldman SR, Fleischer AB, et al. Incidence estimate of nonmelanoma skin cancer in the United States, 2006. Archives of dermatology 2010;146:283–7 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
