

Ensemble modeling highlights importance of understanding parasite-host behavior in preclinical antimalarial drug development
- PMID: 32157151
- PMCID: PMC7064600
- DOI: 10.1038/s41598-020-61304-8
Ensemble modeling highlights importance of understanding parasite-host behavior in preclinical antimalarial drug development
Abstract
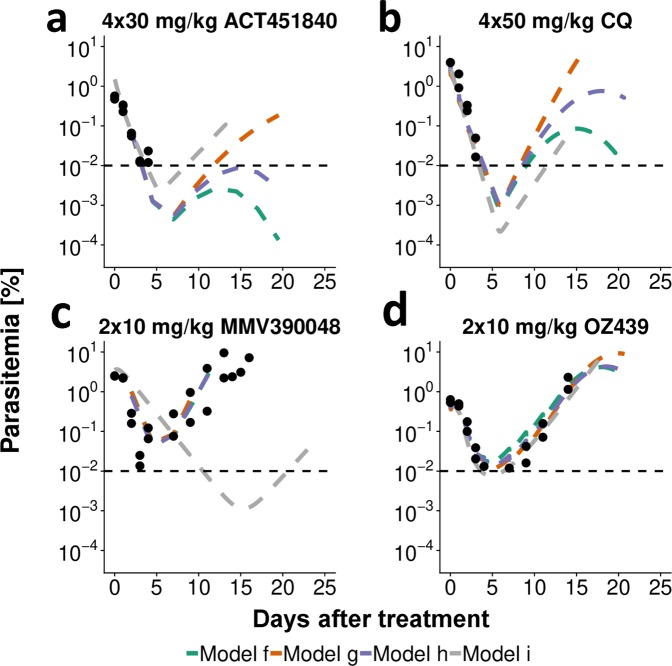
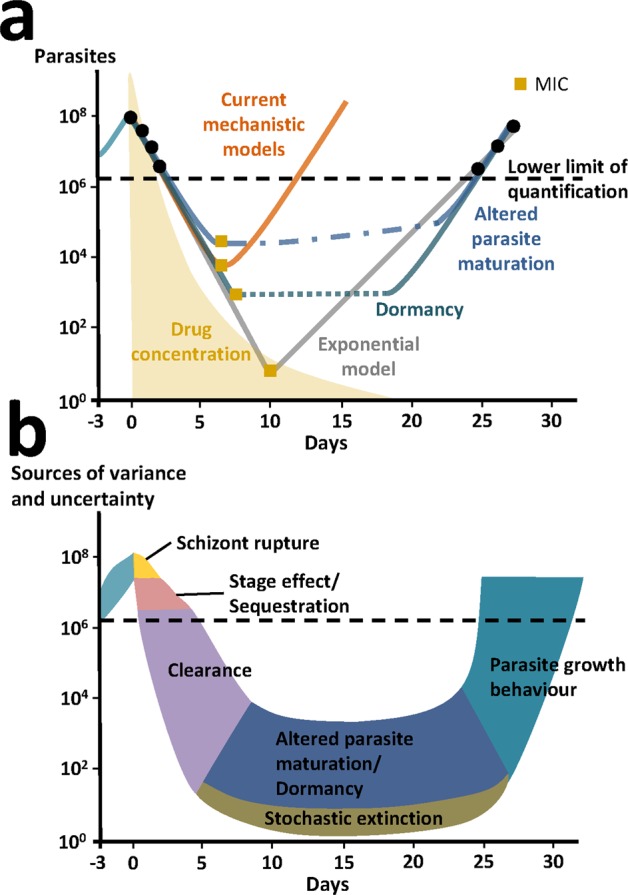
Emerging drug resistance and high-attrition rates in early and late stage drug development necessitate accelerated development of antimalarial compounds. However, systematic and meaningful translation of drug efficacy and host-parasite dynamics between preclinical testing stages is missing. We developed an ensemble of mathematical within-host parasite growth and antimalarial action models, fitted to extensive data from four antimalarials with different modes of action, to assess host-parasite interactions in two preclinical drug testing systems of murine parasite P. berghei in mice, and human parasite P. falciparum in immune-deficient mice. We find properties of the host-parasite system, namely resource availability, parasite maturation and virulence, drive P. berghei dynamics and drug efficacy, whereas experimental constraints primarily influence P. falciparum infection and drug efficacy. Furthermore, uninvestigated parasite behavior such as dormancy influences parasite recrudescence following non-curative treatment and requires further investigation. Taken together, host-parasite interactions should be considered for meaningful translation of pharmacodynamic properties between murine systems and for predicting human efficacious treatment.
Conflict of interest statement
N.G. and J.J.M. are employees of Medicines for Malaria Venture. A.K. and J.D. are employees of Idorsia. All other authors declare no competing interests.
Figures
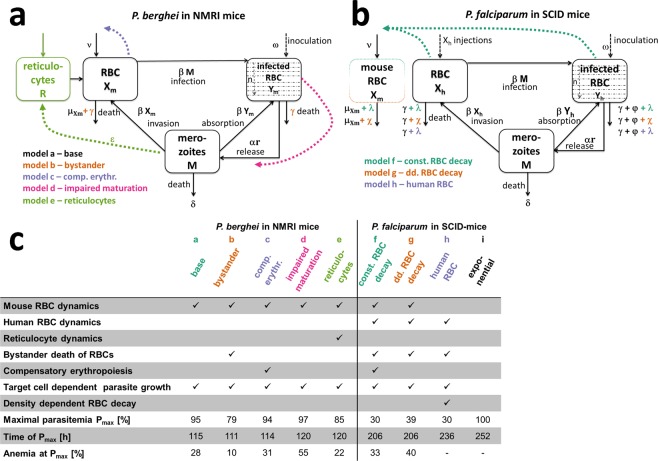
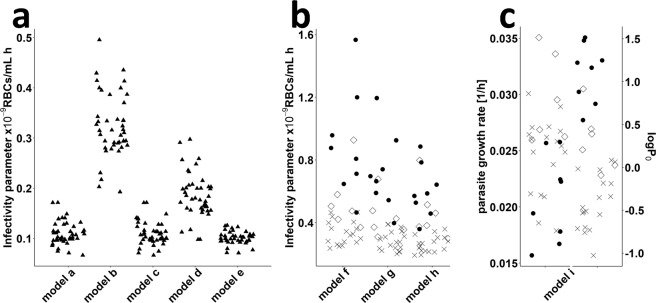
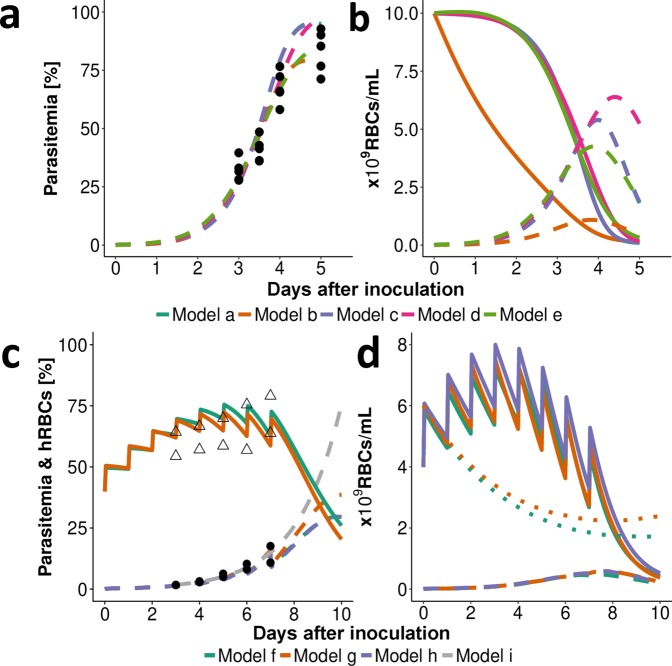
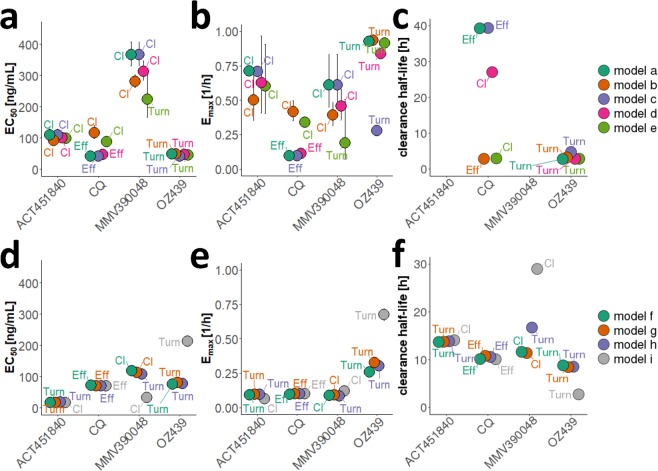
References
-
- World Health Organization. (2018) World malaria report 2018. World Helath Organization. https://apps.who.int/iris/handle/10665/275867. License: CC BY-NC-SA 3.0 IGO
-
- World Health Organization. (2017). Artemisinin and artemisinin-based combination therapy resistance: status report. World Health Organization. https://apps.who.int/iris/handle/10665/255213. License: CC BY-NC-SA 3.0 IGO
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
