

PinX1t, a Novel PinX1 Transcript Variant, Positively Regulates Cardiogenesis of Embryonic Stem Cells
- PMID: 32157956
- PMCID: PMC7335523
- DOI: 10.1161/JAHA.118.010240
PinX1t, a Novel PinX1 Transcript Variant, Positively Regulates Cardiogenesis of Embryonic Stem Cells
Abstract
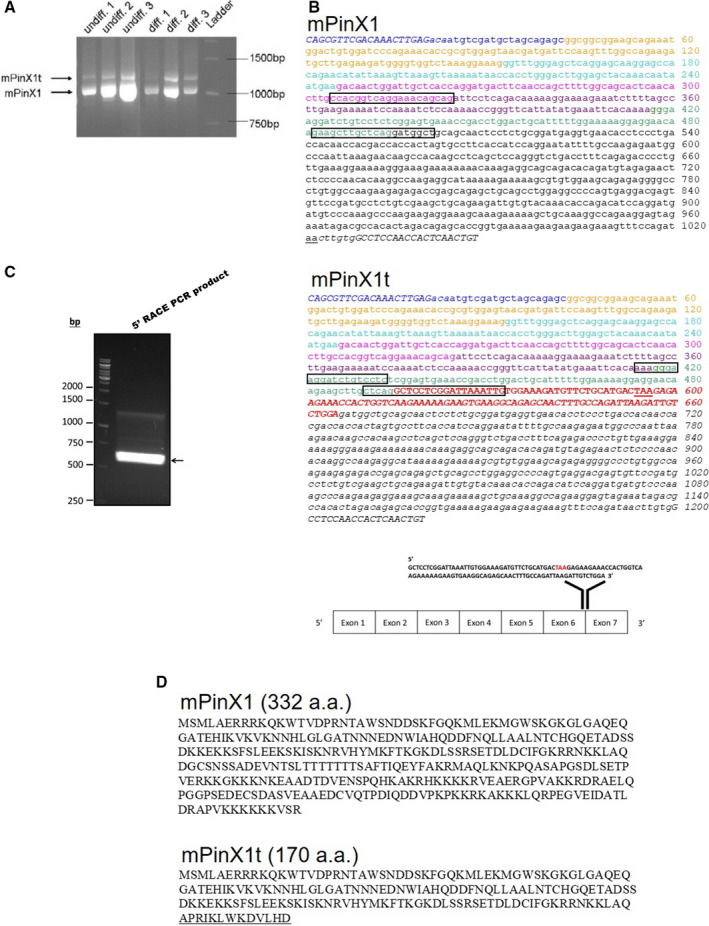
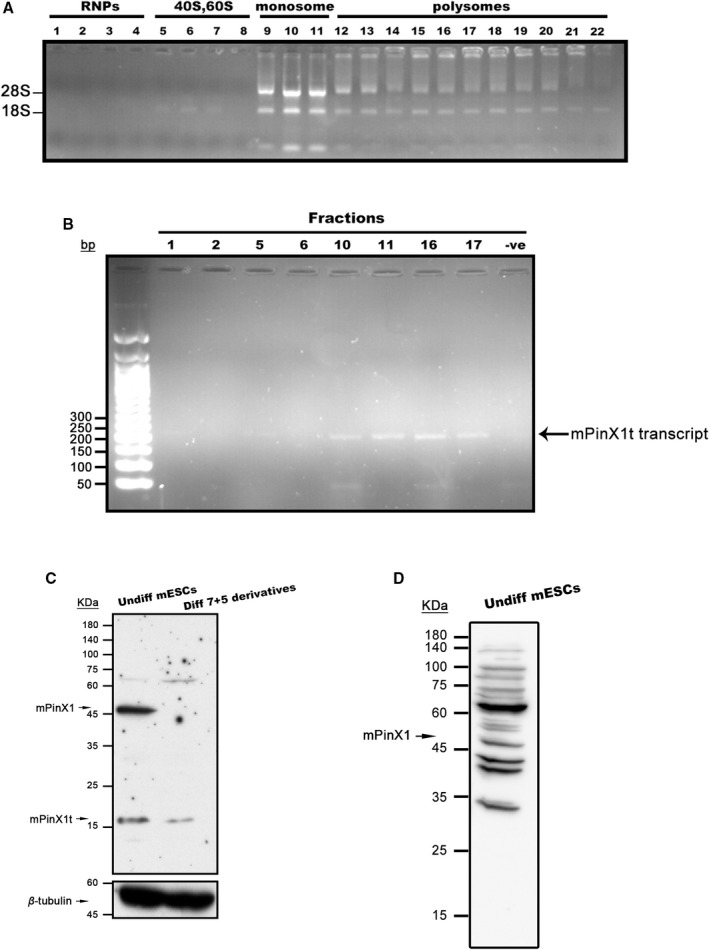
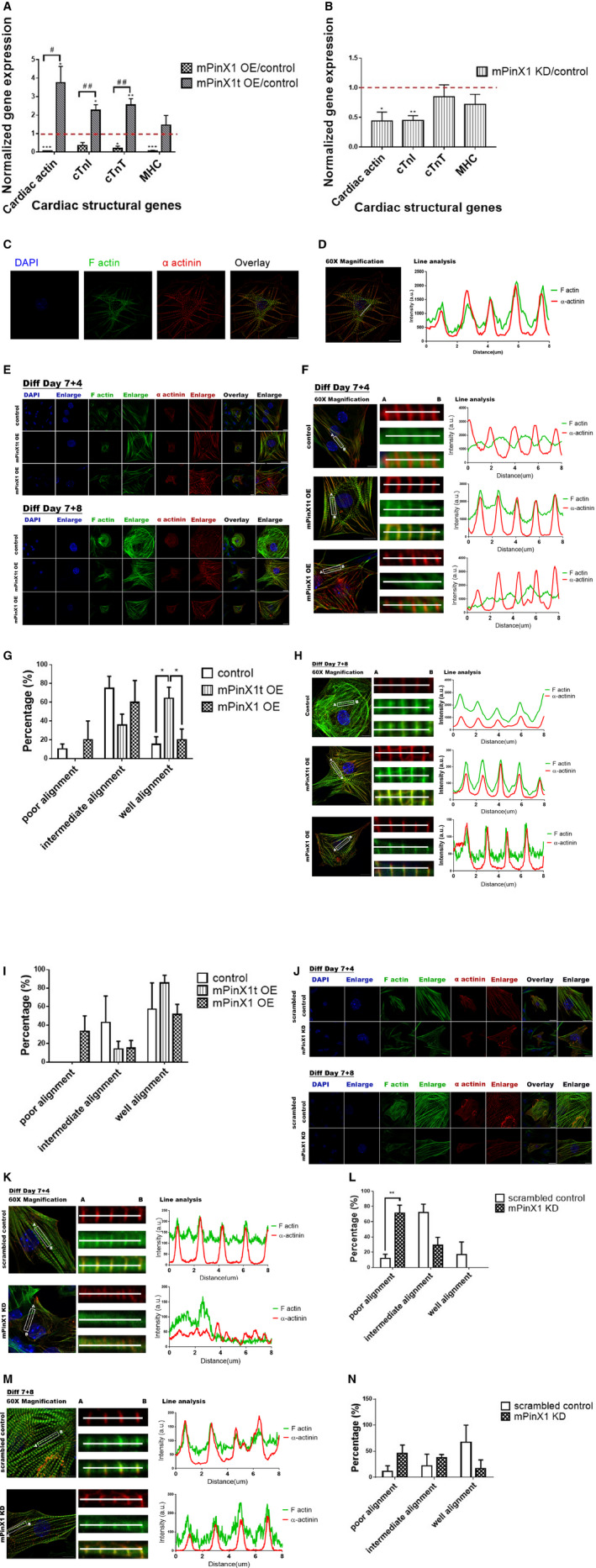
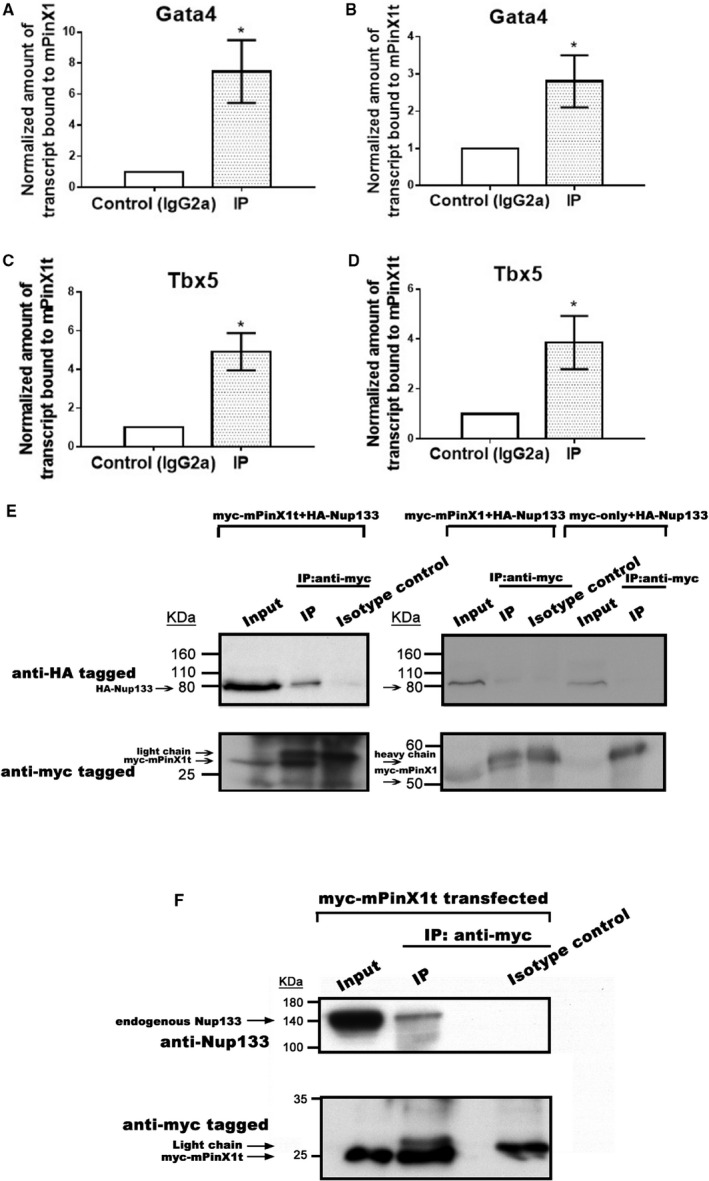
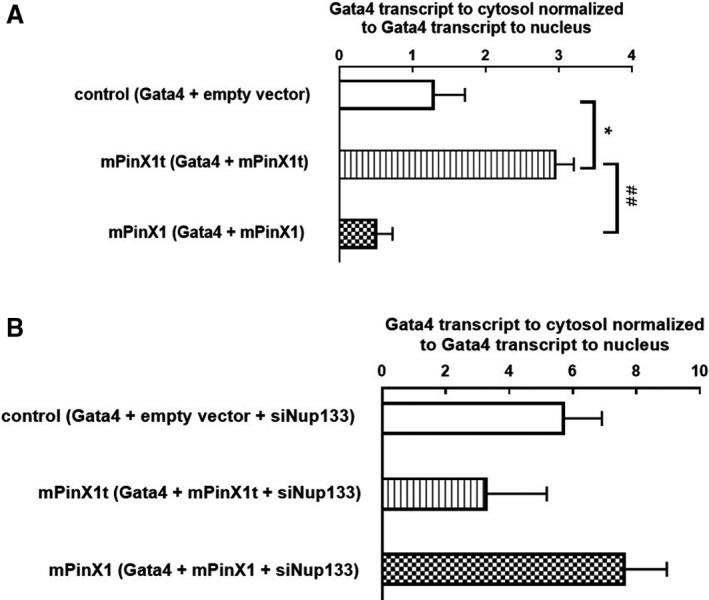
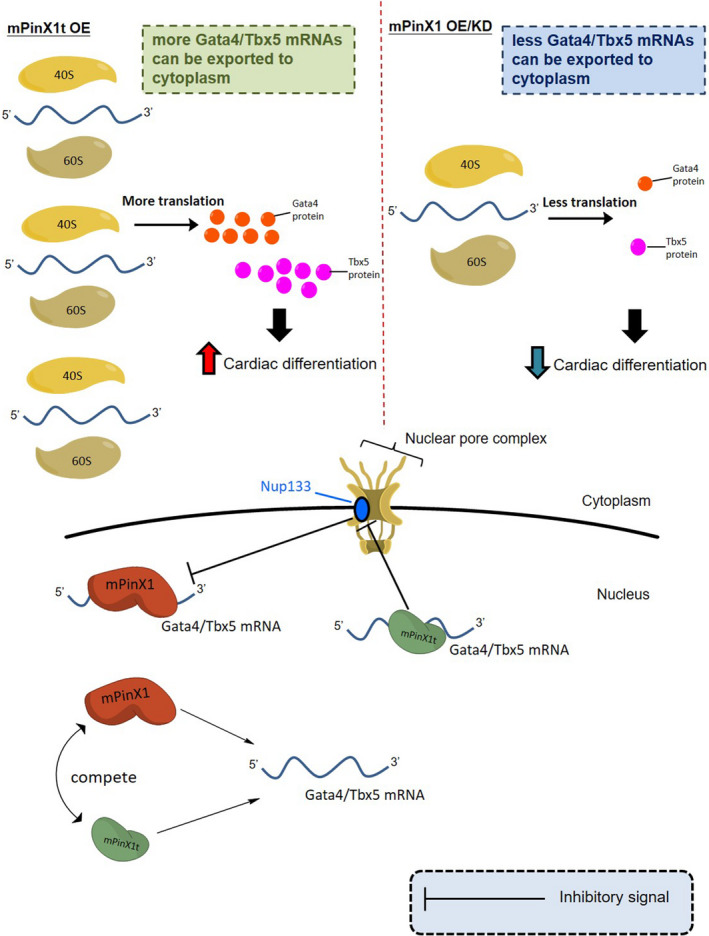
Background Pin2/TRF1-interacting protein, PinX1, was previously identified as a tumor suppressor. Here, we discovered a novel transcript variant of mPinX1 (mouse PinX1), mPinX1t (mouse PinX1t), in embryonic stem cells (ESCs). The aims of this investigation were (1) to detect the presence of mPinX1 and mPinX1t in ESCs and their differentiation derivatives; (2) to investigate the role of mPinX1 and mPinX1t on regulating the characteristics of undifferentiated ESCs and the cardiac differentiation of ESCs; (3) to elucidate the molecular mechanisms of how mPinX1 and mPinX1t regulate the cardiac differentiation of ESCs. Methods and Results By 5' rapid amplification of cDNA ends, 3' rapid amplification of cDNA ends, and polysome fractionation followed by reverse transcription-polymerase chain reaction, mPinX1t transcript was confirmed to be an intact mRNA that is actively translated. Western blot confirmed the existence of mPinX1t protein. Overexpression or knockdown of mPinX1 (both decreased mPinX1t expression) both decreased while overexpression of mPinX1t increased the cardiac differentiation of ESCs. Although both mPinX1 and mPinX1t proteins were found to bind to cardiac transcription factor mRNAs, only mPinX1t protein but not mPinX1 protein was found to bind to nucleoporin 133 protein, a nuclear pore complex component. In addition, mPinX1t-containing cells were found to have a higher cytosol-to-nucleus ratio of cardiac transcription factor mRNAs when compared with that in the control cells. Our data suggested that mPinX1t may positively regulate cardiac differentiation by enhancing export of cardiac transcription factor mRNAs through interacting with nucleoporin 133. Conclusions We discovered a novel transcript variant of mPinX1, the mPinX1t, which positively regulates the cardiac differentiation of ESCs.
Keywords: PinX1; cardiac development; cardiac differentiation; embryonic stem cell; transcript variants.
Figures
References
-
- Scott IC. Life before Nkx2. 5: cardiovascular progenitor cells: embryonic origins and development. Curr Top Dev Biol. 2012;100:1–31. - PubMed
-
- Bruneau BG, Logan M, Davis N, Levi T, Tabin CJ, Seidman JG, Seidman CE. Chamber‐specific cardiac expression of Tbx5 and heart defects in Holt‐Oram syndrome. Dev Biol. 1999;211:100–108. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
