

HIF-1 Has a Central Role in Caenorhabditis elegans Organismal Response to Selenium
- PMID: 32161616
- PMCID: PMC7052493
- DOI: 10.3389/fgene.2020.00063
HIF-1 Has a Central Role in Caenorhabditis elegans Organismal Response to Selenium
Abstract
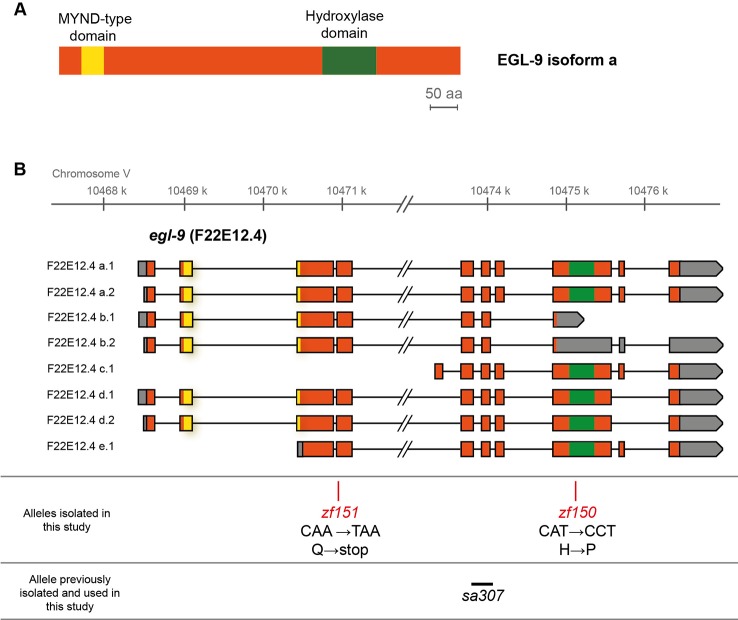
Selenium is a trace element for most organisms; its deficiency and excess are detrimental. Selenium beneficial effects are mainly due to the role of the 21st genetically encoded amino acid selenocysteine (Sec). Selenium also exerts Sec-independent beneficial effects. Its harmful effects are thought to be mainly due to non-specific incorporation in protein synthesis. Yet the selenium response in animals is poorly understood. In Caenorhabditis elegans, Sec is genetically incorporated into a single selenoprotein. Similar to mammals, a 20-fold excess of the optimal selenium requirement is harmful. Sodium selenite (Na2SeO3) excess causes development retardation, impaired growth, and neurodegeneration of motor neurons. To study the organismal response to selenium we performed a genetic screen for C. elegans mutants that are resistant to selenite. We isolated non-sense and missense egl-9/EGLN mutants that confer robust resistance to selenium. In contrast, hif-1/HIF null mutant was highly sensitive to selenium, establishing a role for this transcription factor in the selenium response. We showed that EGL-9 regulates HIF-1 activity through VHL-1, and identified CYSL-1 as a key sensor that transduces the selenium signal. Finally, we showed that the key enzymes involved in sulfide and sulfite stress (sulfide quinone oxidoreductase and sulfite oxidase) are not required for selenium resistance. In contrast, knockout strains in the persulfide dioxygenase ETHE-1 and the sulfurtransferase MPST-7 affect the organismal response to selenium. In sum, our results identified a transcriptional pathway as well as enzymes possibly involved in the organismal selenium response.
Keywords: CYSL-1; Caenorhabditis elegans; EGL-9; HIF-1; selenite; selenium; stress; sulfide.
Copyright © 2020 Romanelli-Credrez, Doitsidou, Alkema and Salinas.
Figures




References
-
- Bjornstedt M., Kumar S. (1992). Selenodiglutathione is a highly efficient oxidant of reduced thioredoxin and a substrate for mammalian thioredoxin reductase*. J. Biol. Chem. 267, 8030–8035. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials