

Claudins in Renal Physiology and Pathology
- PMID: 32164158
- PMCID: PMC7140793
- DOI: 10.3390/genes11030290
Claudins in Renal Physiology and Pathology
Abstract
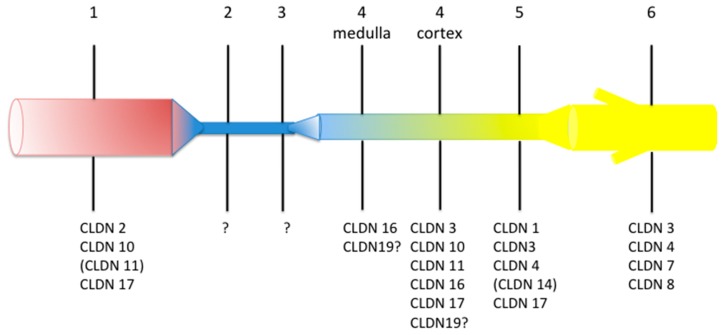
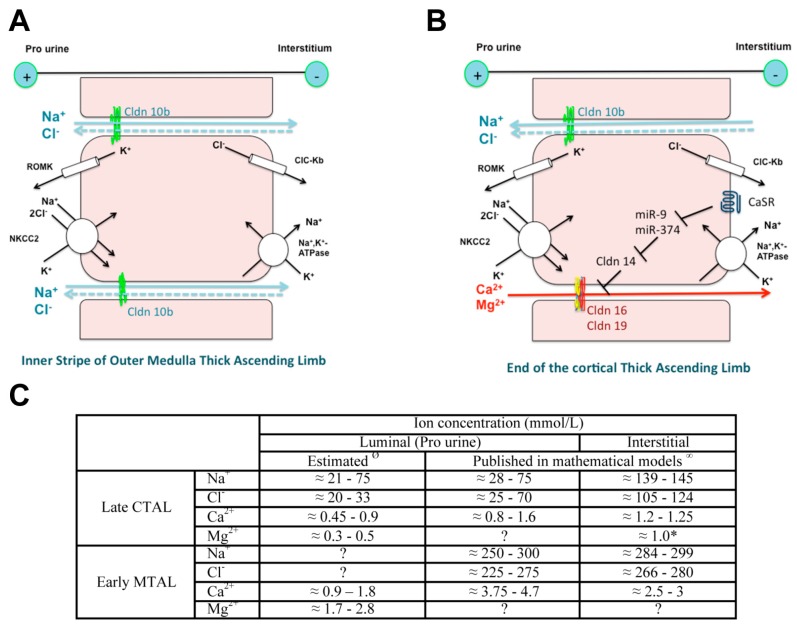
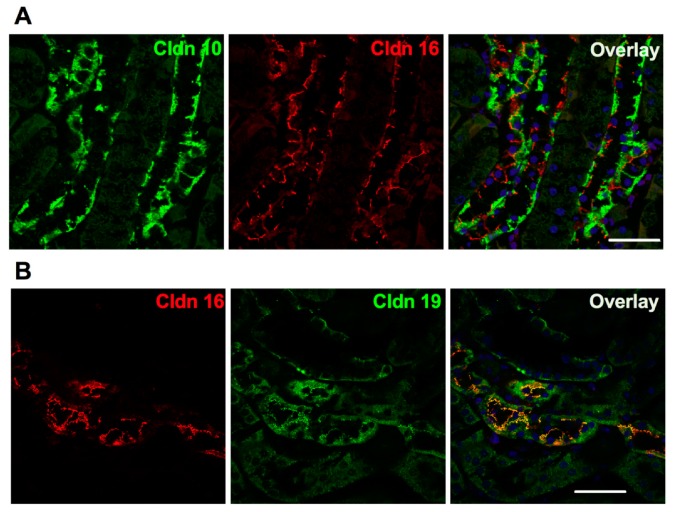
Claudins are integral proteins expressed at the tight junctions of epithelial and endothelial cells. In the mammalian kidney, every tubular segment express a specific set of claudins that give to that segment unique properties regarding permeability and selectivity of the paracellular pathway. So far, 3 claudins (10b, 16 and 19) have been causally traced to rare human syndromes: variants of CLDN10b cause HELIX syndrome and variants of CLDN16 or CLDN19 cause familial hypomagnesemia with hypercalciuria and nephrocalcinosis. The review summarizes our current knowledge on the physiology of mammalian tight junctions and paracellular ion transport, as well as on the role of the 3 above-mentioned claudins in health and disease. Claudin 14, although not having been causally linked to any rare renal disease, is also considered, because available evidence suggests that it may interact with claudin 16. Some single-nucleotide polymorphisms of CLDN14 are associated with urinary calcium excretion and/or kidney stones. For each claudin considered, the pattern of expression, the function and the human syndrome caused by pathogenic variants are described.
Keywords: HELIX syndrome; claudin 10b; claudin 14; claudin 16; claudin 19; divalent cations; familial hypomagnesemia with hypercalciuria and nephrocalcinosis; kidney; sodium; tight junction.
Conflict of interest statement
C.P.-B. and P.H. have no conflict of interest to declare regarding the content of the present review.
Figures
References
-
- Reuss L. Tight junction permeability to ions and water. In: Cereijido M., Anderson J.M., editors. Tight Junctions. 2nd ed. CRC Press; Boca Raton, FL, USA: 2001. pp. 61–88.
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
