

The tethering function of mitofusin2 controls osteoclast differentiation by modulating the Ca2+-NFATc1 axis
- PMID: 32165499
- PMCID: PMC7212632
- DOI: 10.1074/jbc.RA119.012023
The tethering function of mitofusin2 controls osteoclast differentiation by modulating the Ca2+-NFATc1 axis
Abstract
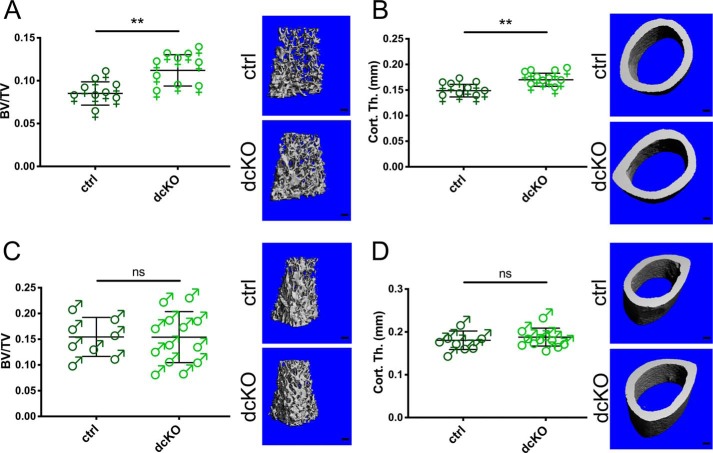
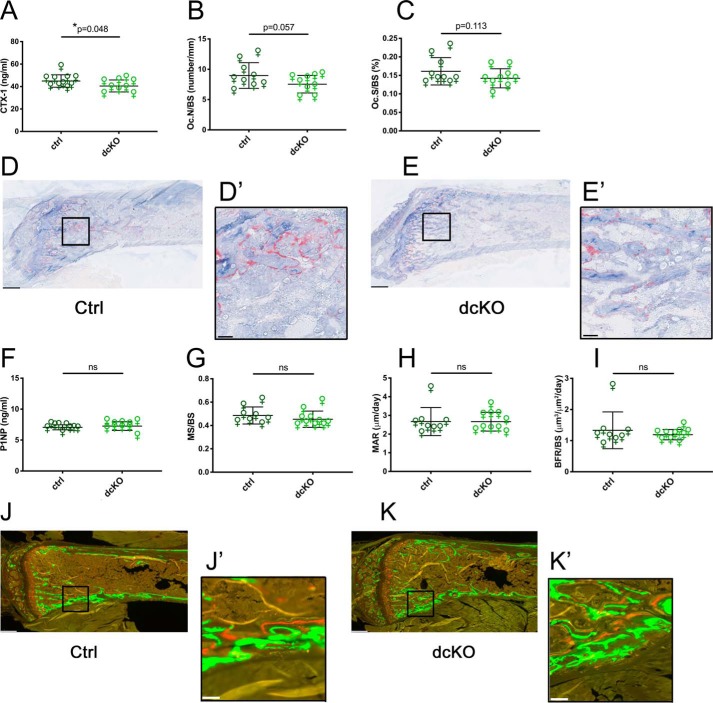
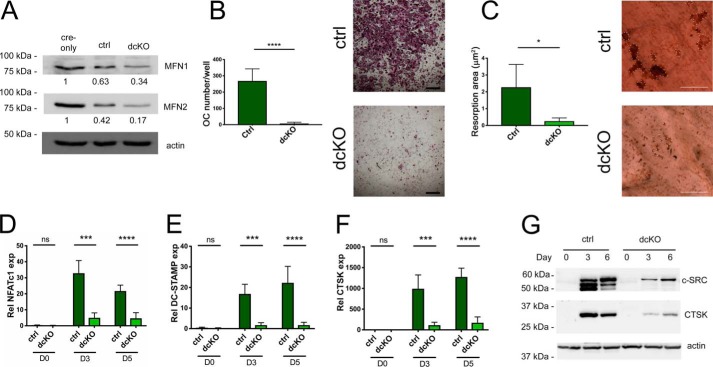
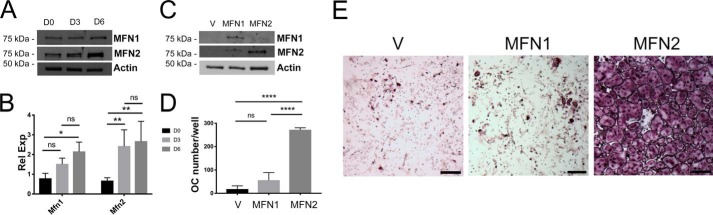
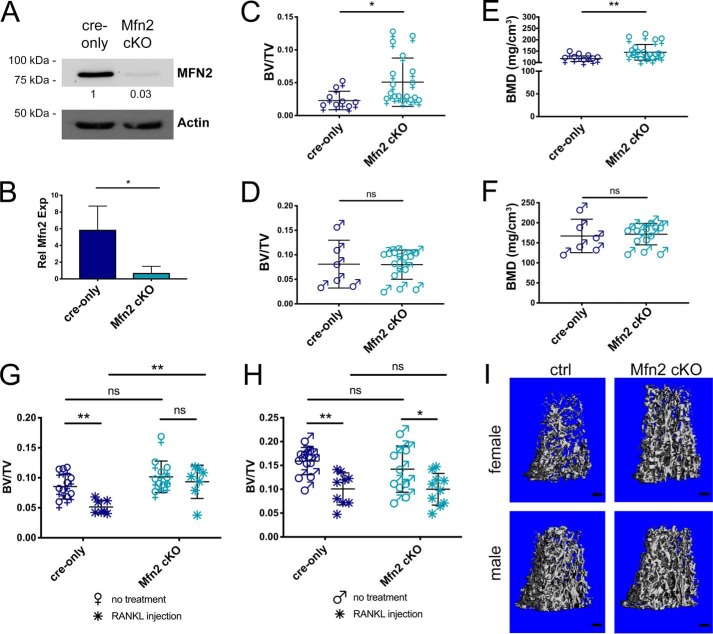
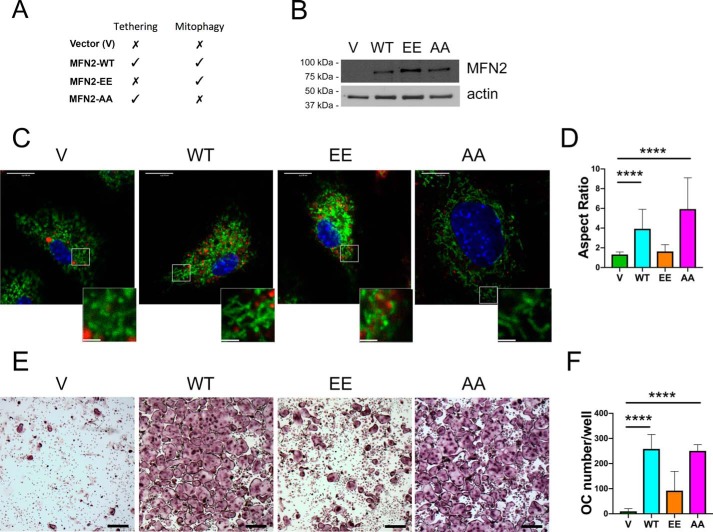
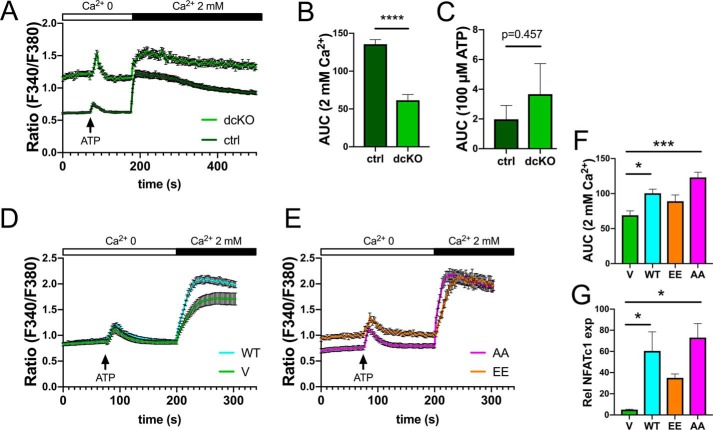
Dynamic regulation of the mitochondrial network by mitofusins (MFNs) modulates energy production, cell survival, and many intracellular signaling events, including calcium handling. However, the relative importance of specific mitochondrial functions and their dependence on MFNs vary greatly among cell types. Osteoclasts have many mitochondria, and increased mitochondrial biogenesis and oxidative phosphorylation enhance bone resorption, but little is known about the mitochondrial network or MFNs in osteoclasts. Because expression of each MFN isoform increases with osteoclastogenesis, we conditionally deleted MFN1 and MFN2 (double conditional KO (dcKO)) in murine osteoclast precursors, finding that this increased bone mass in young female mice and abolished osteoclast precursor differentiation into mature osteoclasts in vitro Defective osteoclastogenesis was reversed by overexpression of MFN2 but not MFN1; therefore, we generated mice lacking only MFN2 in osteoclasts. MFN2-deficient female mice had increased bone mass at 1 year and resistance to Receptor Activator of NF-κB Ligand (RANKL)-induced osteolysis at 8 weeks. To explore whether MFN-mediated tethering or mitophagy is important for osteoclastogenesis, we overexpressed MFN2 variants defective in either function in dcKO precursors and found that, although mitophagy was dispensable for differentiation, tethering was required. Because the master osteoclastogenic transcriptional regulator nuclear factor of activated T cells 1 (NFATc1) is calcium-regulated, we assessed calcium release from the endoplasmic reticulum and store-operated calcium entry and found that the latter was blunted in dcKO cells. Restored osteoclast differentiation by expression of intact MFN2 or the mitophagy-defective variant was associated with normalization of store-operated calcium entry and NFATc1 levels, indicating that MFN2 controls mitochondrion-endoplasmic reticulum tethering in osteoclasts.
Keywords: Mus musculus; animal model; bone; calcium; mitochondria; mitochondrial dynamics; osteoclast; osteoclastogenesis.
© 2020 Ballard et al.
Conflict of interest statement
G. W. D. is a founder of Mitochondria in Motion, Inc., a St. Louis–based startup biotech R&D company focused on enhancing mitochondrial trafficking and fitness in neurodegenerative diseases, and may financially benefit if the company is successful in marketing products related to this research. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
