

Regulation of keratin network dynamics by the mechanical properties of the environment in migrating cells
- PMID: 32165652
- PMCID: PMC7067805
- DOI: 10.1038/s41598-020-61242-5
Regulation of keratin network dynamics by the mechanical properties of the environment in migrating cells
Abstract
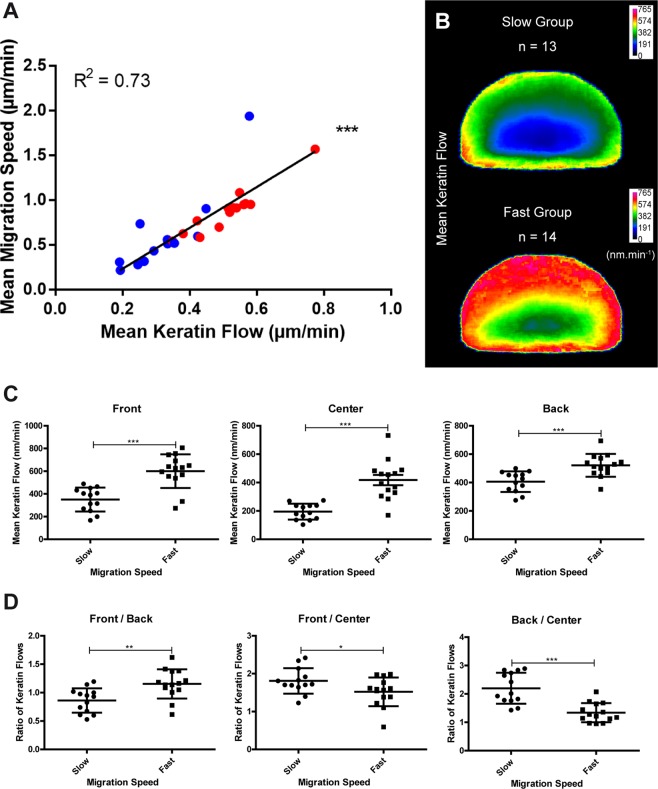
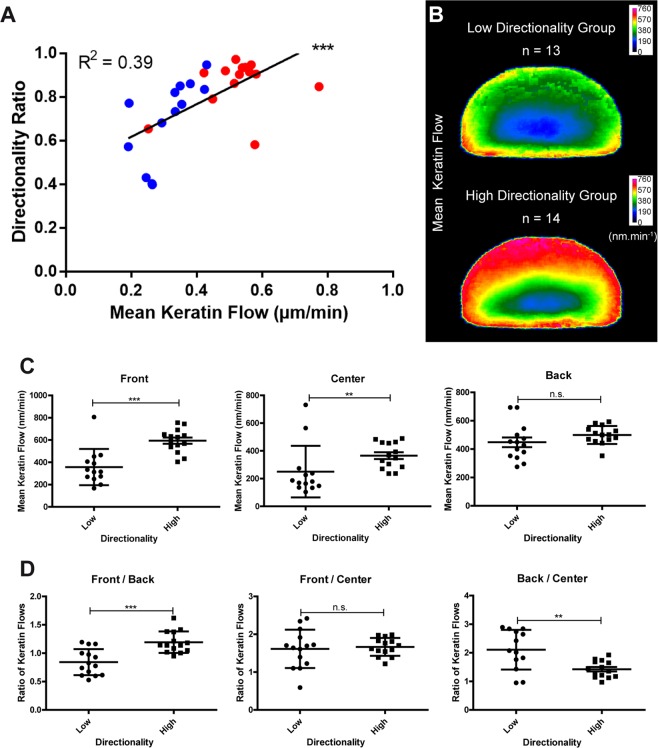
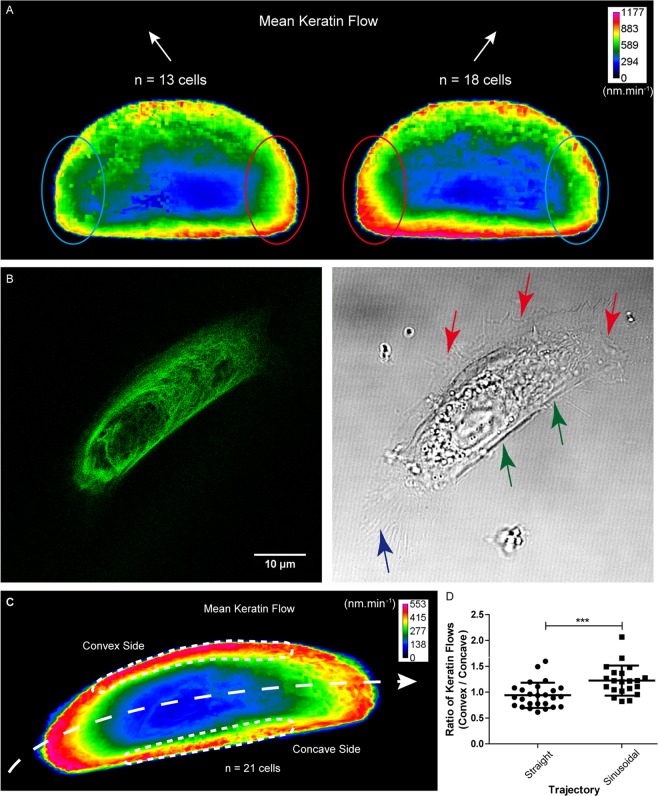
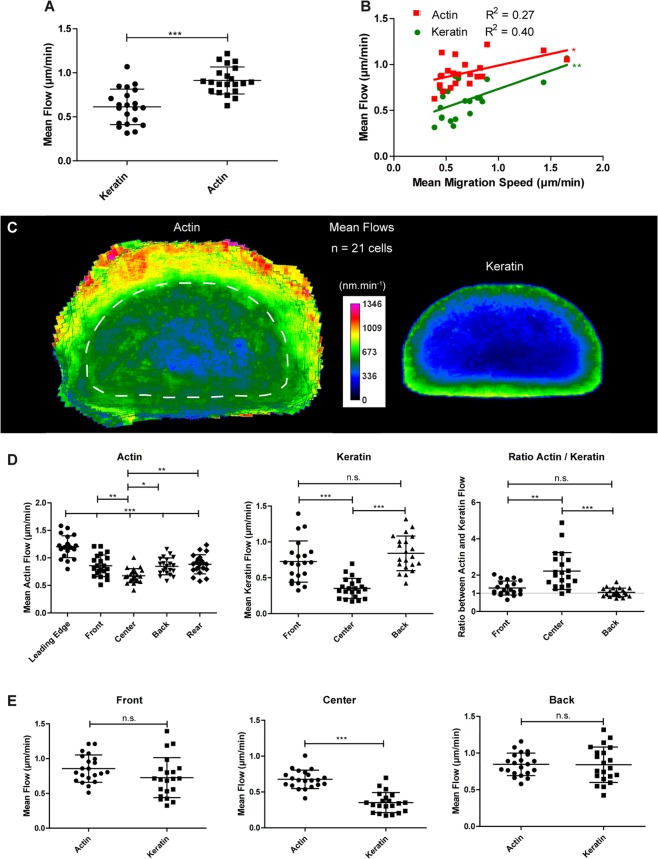
Keratin intermediate filaments provide mechanical resilience for epithelia. They are nevertheless highly dynamic and turn over continuously, even in sessile keratinocytes. The aim of this study was to characterize and understand how the dynamic behavior of the keratin cytoskeleton is integrated in migrating cells. By imaging human primary keratinocytes producing fluorescent reporters and by using standardized image analysis we detect inward-directed keratin flow with highest rates in the cell periphery. The keratin flow correlates with speed and trajectory of migration. Changes in fibronectin-coating density and substrate stiffness induces concordant changes in migration speed and keratin flow. When keratinocytes are pseudo-confined on stripes, migration speed and keratin flow are reduced affecting the latter disproportionately. The regulation of keratin flow is linked to the regulation of actin flow. Local speed and direction of keratin and actin flow are very similar in migrating keratinocytes with keratin flow lagging behind actin flow. Conversely, reduced actin flow in areas of high keratin density indicates an inhibitory function of keratins on actin dynamics. Together, we propose that keratins enhance persistence of migration by directing actin dynamics and that the interplay of keratin and actin dynamics is modulated by matrix adhesions.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
