

Predation risk shapes the degree of placentation in natural populations of live-bearing fish
- PMID: 32166847
- PMCID: PMC7187176
- DOI: 10.1111/ele.13487
Predation risk shapes the degree of placentation in natural populations of live-bearing fish
Abstract
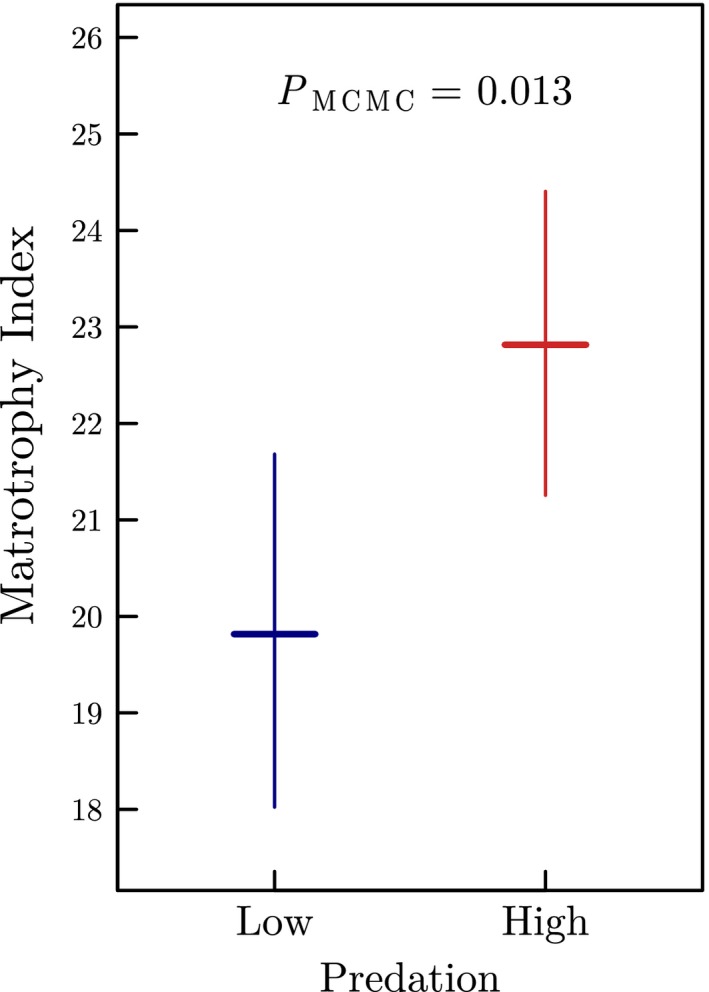
The placenta is a complex life-history trait that is ubiquitous across the tree of life. Theory proposes that the placenta evolves in response to high performance-demanding conditions by shifting maternal investment from pre- to post-fertilisation, thereby reducing a female's reproductive burden during pregnancy. We test this hypothesis by studying populations of the fish species Poeciliopsis retropinna in Costa Rica. We found substantial variation in the degree of placentation among natural populations associated with predation risk: females from high predation populations had significantly higher degrees of placentation compared to low predation females, while number, size and quality of offspring at birth remained unaffected. Moreover, a higher degree of placentation correlated with a lower reproductive burden and hence likely an improved swimming performance during pregnancy. Our study advances an adaptive explanation for why the placenta evolves by arguing that an increased degree of placentation offers a selective advantage in high predation environments.
Keywords: Life-history; Poeciliidae; Trexler-DeAngelis; live-bearing; matrotrophy; placenta; placentotrophy; predation; superfetation; viviparity.
© 2020 The Authors. Ecology Letters published by CNRS and John Wiley & Sons Ltd.
Figures


References
-
- Arnold, S.J. , Pfrender, M.E. & Jones, A.G. (2001). The adaptive landscape as a conceptual bridge between micro‐ and macroevolution. Genetica, 112–113, 9–32. - PubMed
-
- Banet, A.I. & Reznick, D.N. (2008). Do placental species abort offspring? Testing an assumption of the Trexler‐DeAngelis model. Functional Ecology, 22, 323–331.
-
- Banet, A.I. , Au, A.G. & Reznick, D.N. (2010). Is mom in charge? Implications of resource provisioning on the evolution of the placenta. Evolution, 64, 3172–3182. - PubMed
-
- Bashey, F. (2006). Cross‐generational environmental effects and the evolution of offspring size in the Trinidadian Guppy Poecilia reticulata . Evolution, 60, 348–361. - PubMed
-
- Basolo, A.L. & Wagner, W.E. (2004). Covariation between predation risk, body size and fin elaboration in the green swordtail, Xiphophorus helleri . Biol. J. Linn. Soc., 83, 87–100.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
