

Initiation of Parental Genome Reprogramming in Fertilized Oocyte by Splicing Kinase SRPK1-Catalyzed Protamine Phosphorylation
- PMID: 32169215
- PMCID: PMC7190278
- DOI: 10.1016/j.cell.2020.02.020
Initiation of Parental Genome Reprogramming in Fertilized Oocyte by Splicing Kinase SRPK1-Catalyzed Protamine Phosphorylation
Abstract
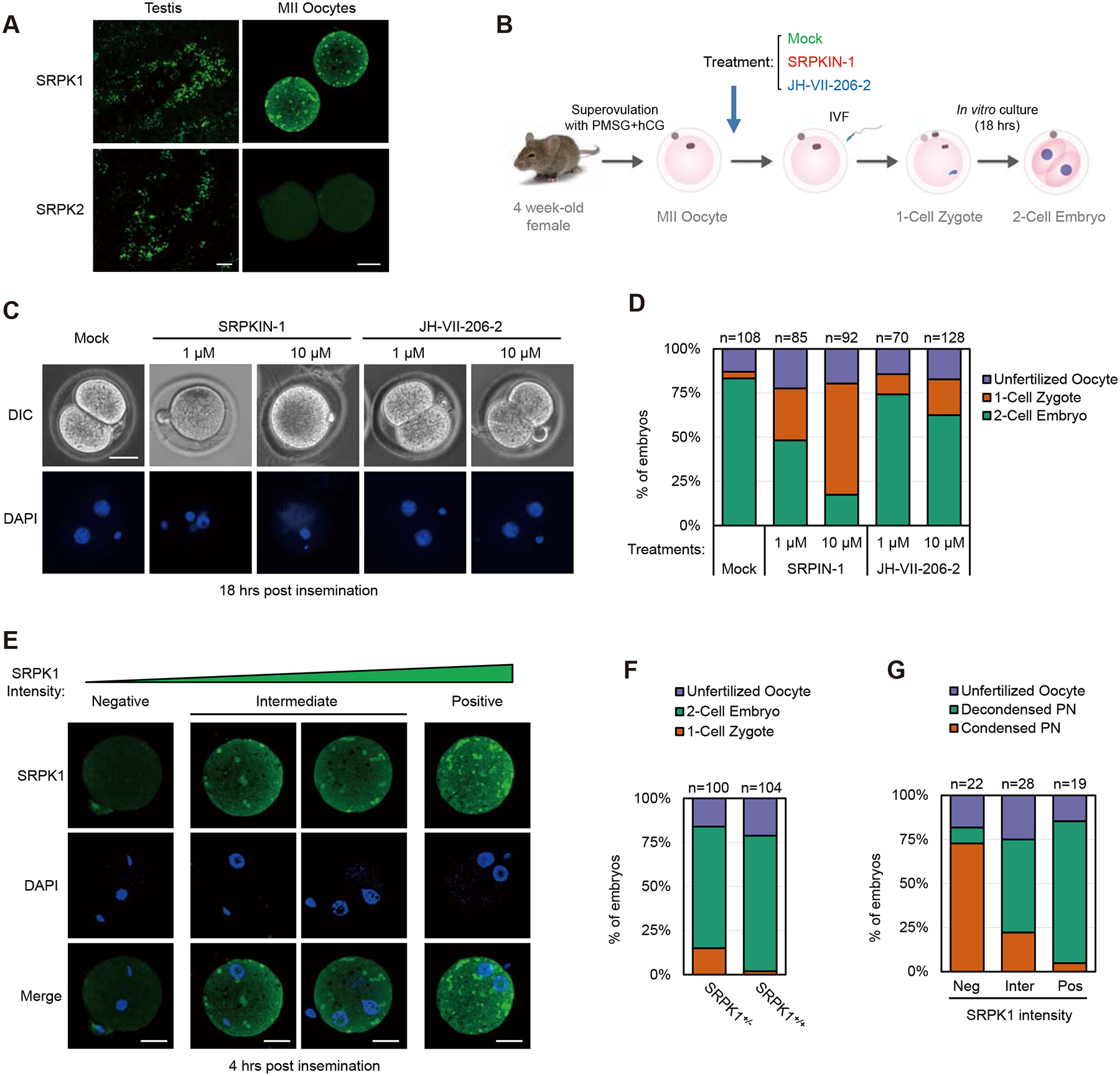
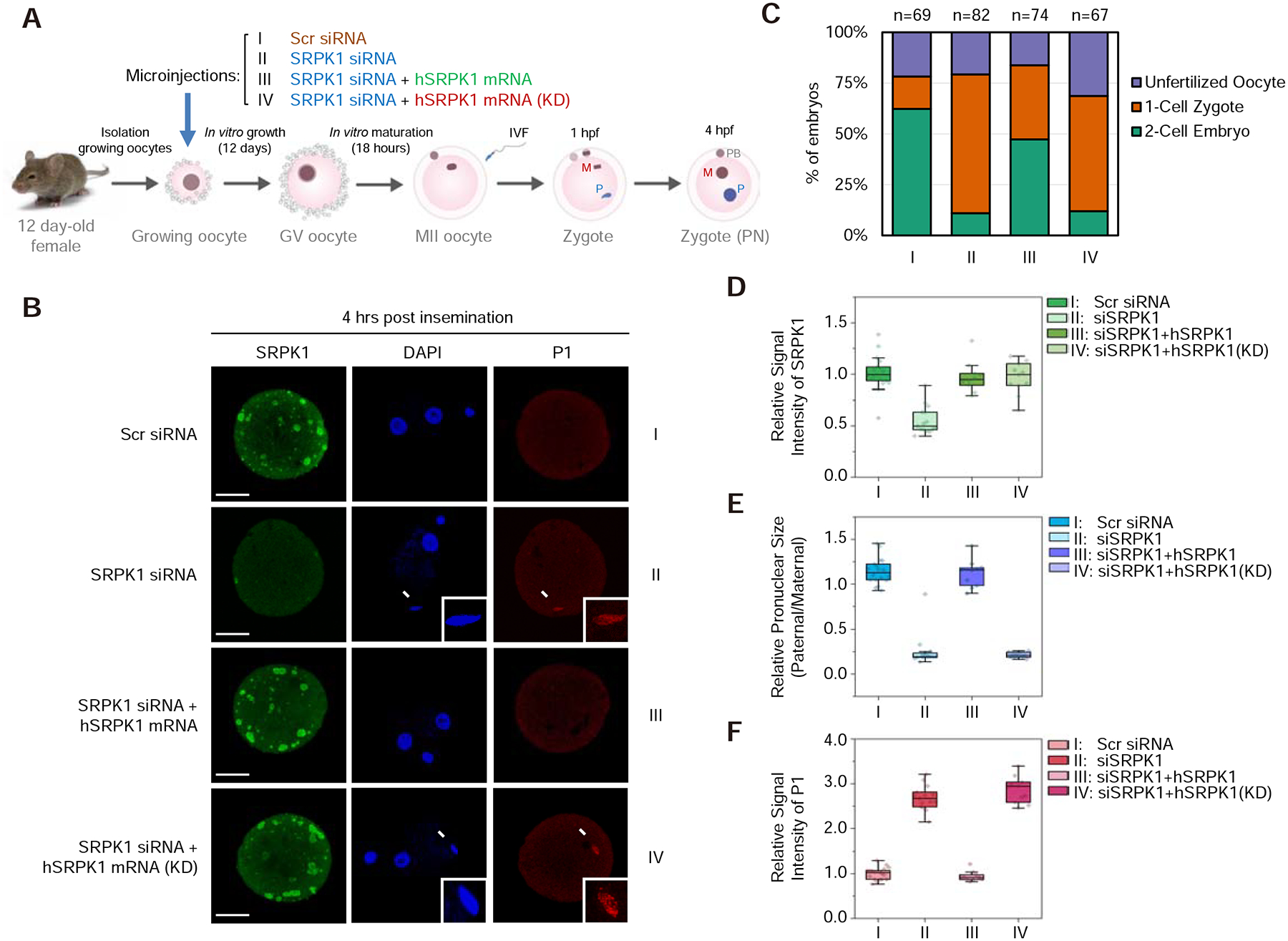
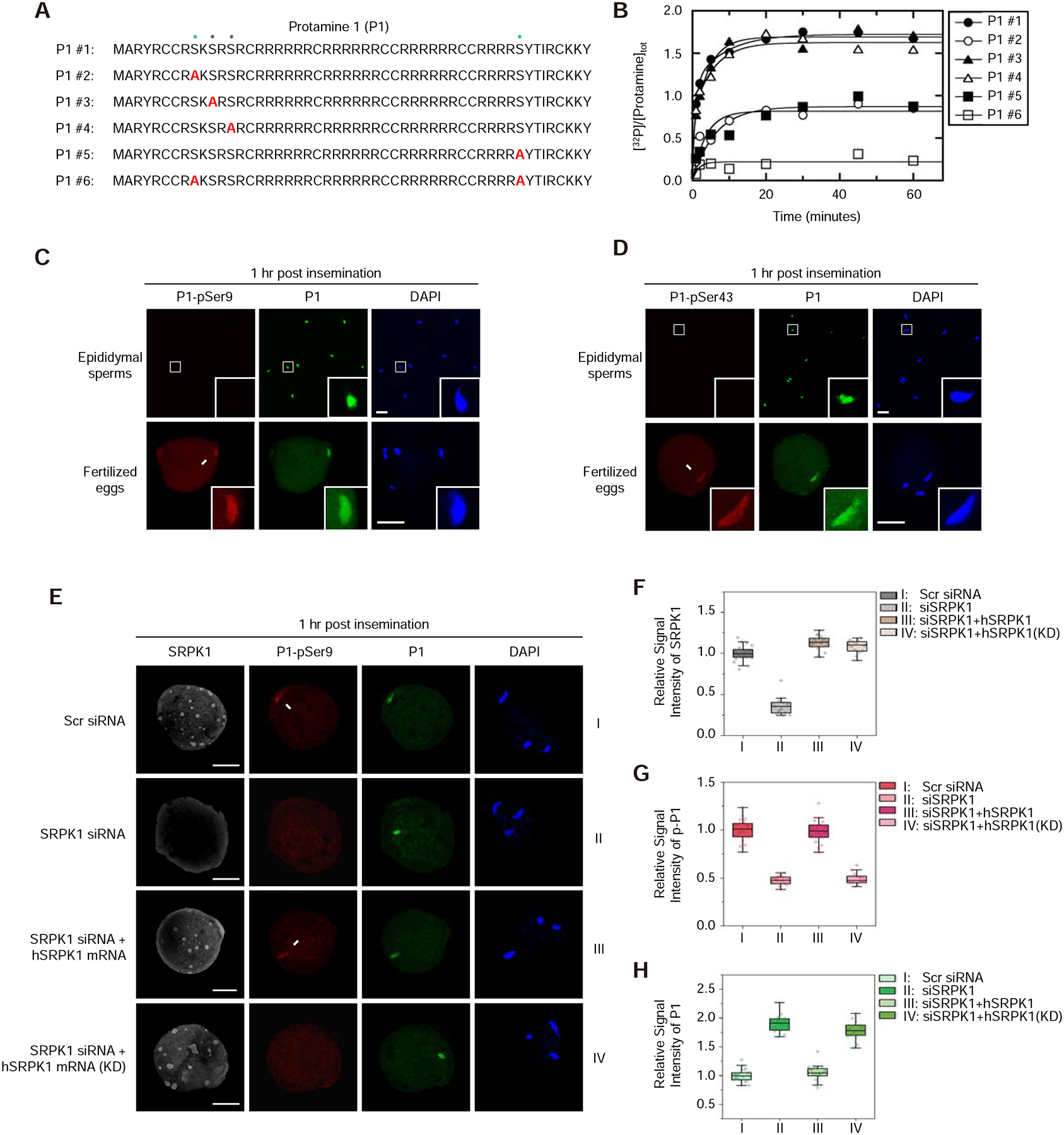
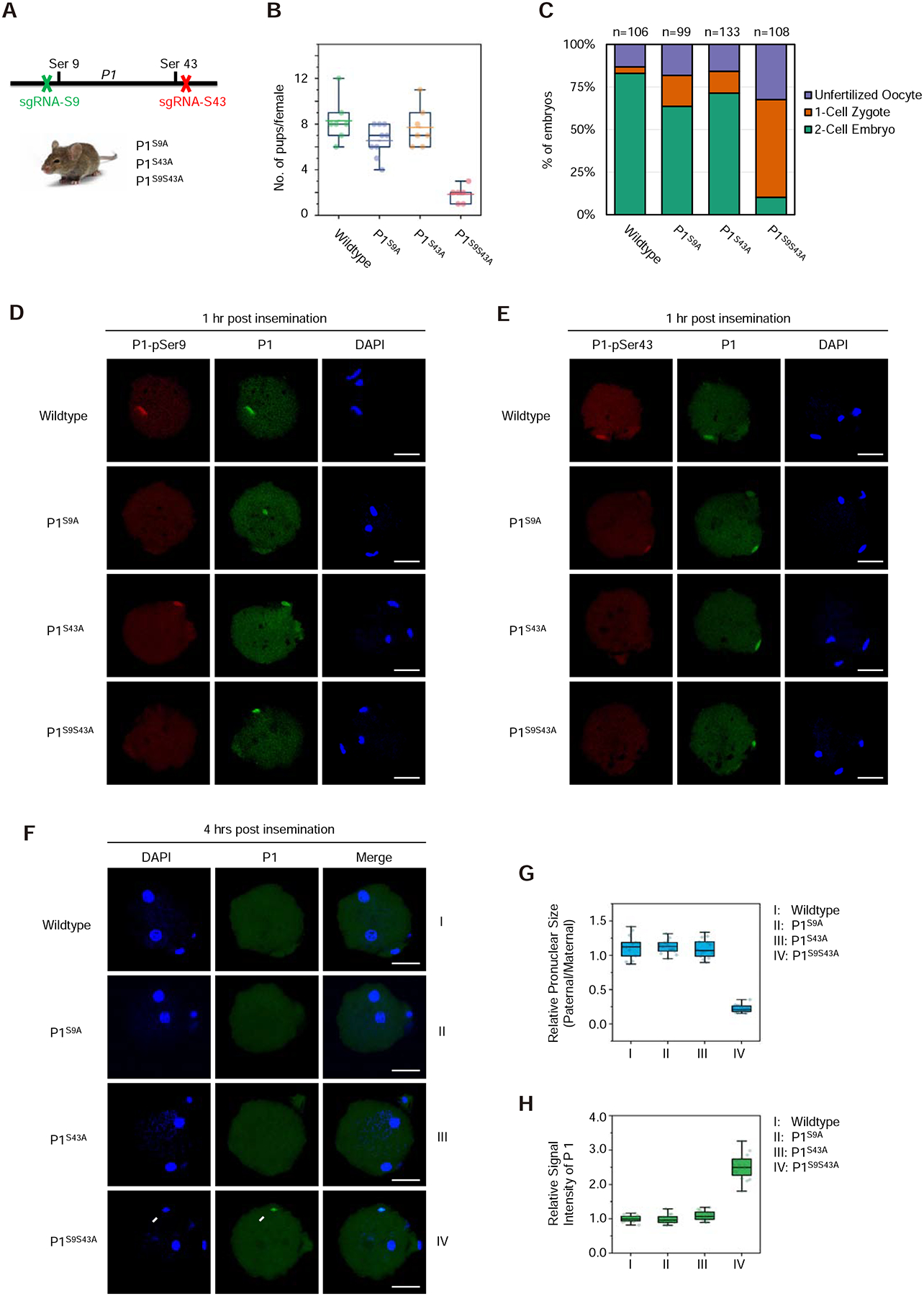
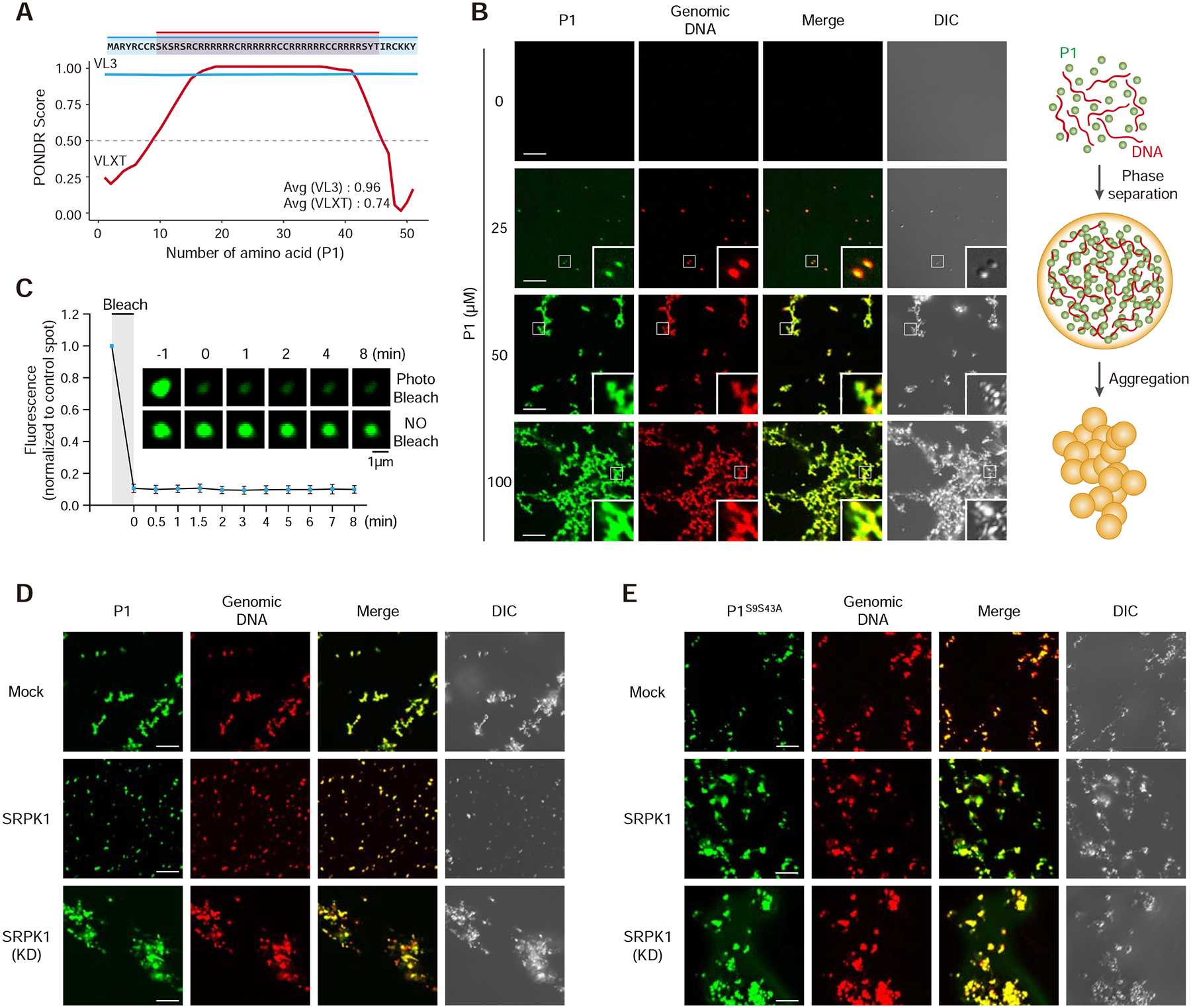
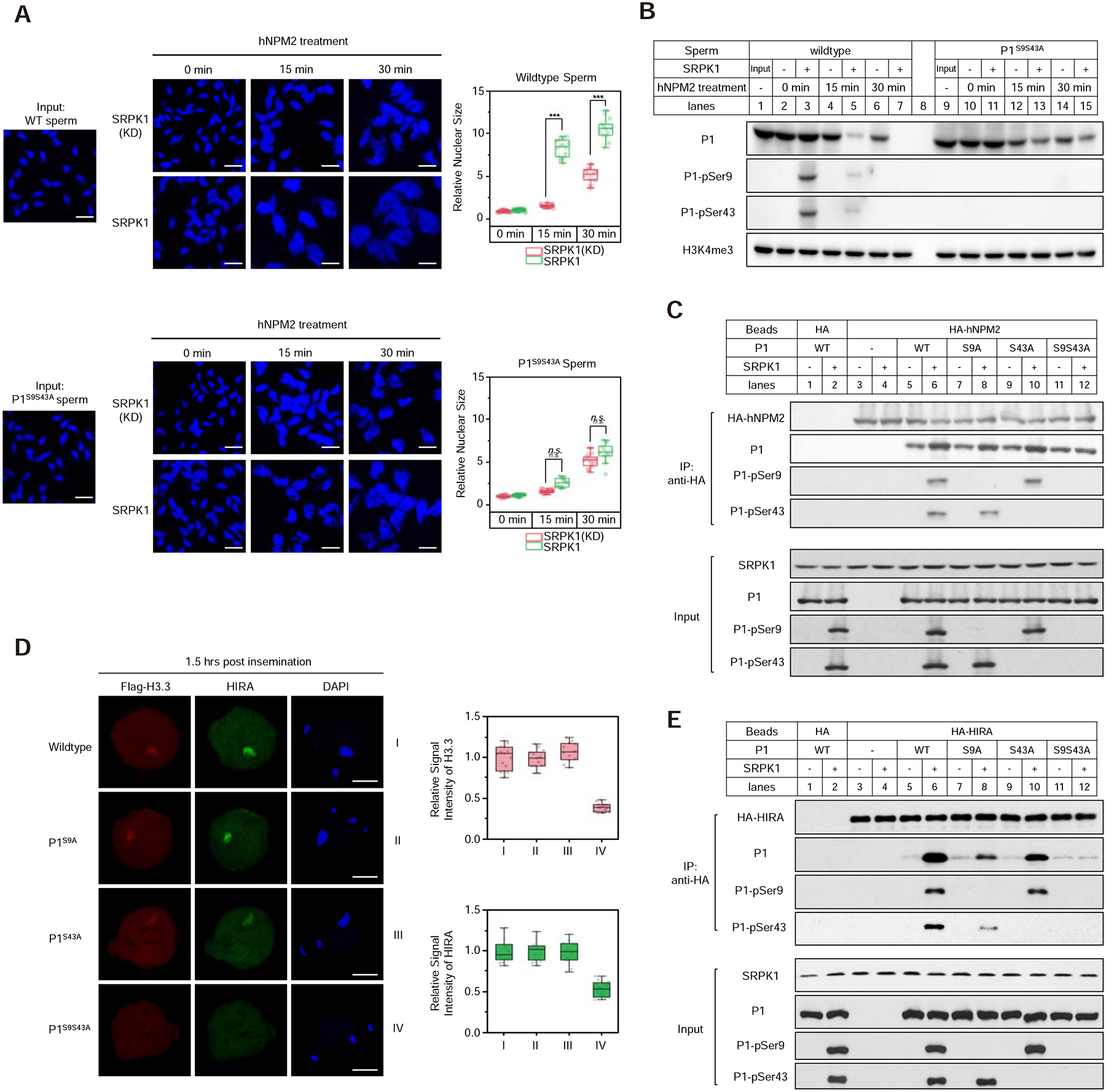
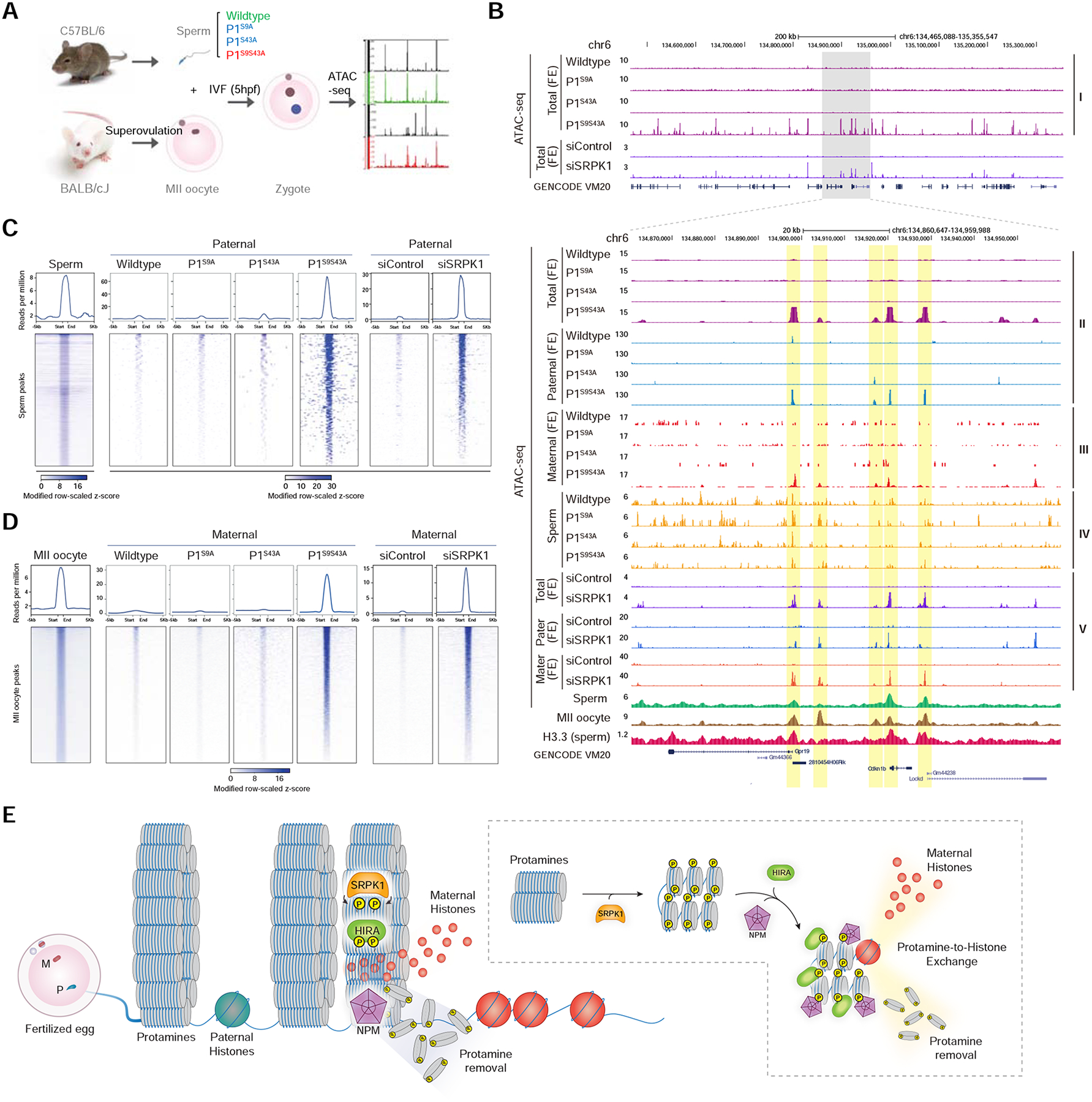
The paternal genome undergoes a massive exchange of histone with protamine for compaction into sperm during spermiogenesis. Upon fertilization, this process is potently reversed, which is essential for parental genome reprogramming and subsequent activation; however, it remains poorly understood how this fundamental process is initiated and regulated. Here, we report that the previously characterized splicing kinase SRPK1 initiates this life-beginning event by catalyzing site-specific phosphorylation of protamine, thereby triggering protamine-to-histone exchange in the fertilized oocyte. Interestingly, protamine undergoes a DNA-dependent phase transition to gel-like condensates and SRPK1-mediated phosphorylation likely helps open up such structures to enhance protamine dismissal by nucleoplasmin (NPM2) and enable the recruitment of HIRA for H3.3 deposition. Remarkably, genome-wide assay for transposase-accessible chromatin sequencing (ATAC-seq) analysis reveals that selective chromatin accessibility in both sperm and MII oocytes is largely erased in early pronuclei in a protamine phosphorylation-dependent manner, suggesting that SRPK1-catalyzed phosphorylation initiates a highly synchronized reorganization program in both parental genomes.
Keywords: SR protein-specific kinase; fertilization; genome reprogramming; histone chaperones; phosphorylation; protamine; protamine-to-histone exchange; zygotic development.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing financial interests.
Figures
References
-
- Adenot PG, Szollosi MS, Geze M, Renard JP, and Debey P (1991). Dynamics of Paternal Chromatin Changes in Live One-Cell Mouse Embryo after Natural Fertilization. Mol Reprod Dev 28, 23–34. - PubMed
-
- Balhorn R (1989) Mammalian Protamines: Structure and Molecular Interactions In: Adolph KW (eds) Molecular Biology of Chromosome Function. Springer, New York, NY
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
