

Red Blood Cell Dysfunction in Critical Illness
- PMID: 32172813
- PMCID: PMC7183574
- DOI: 10.1016/j.ccc.2019.12.008
Red Blood Cell Dysfunction in Critical Illness
Abstract
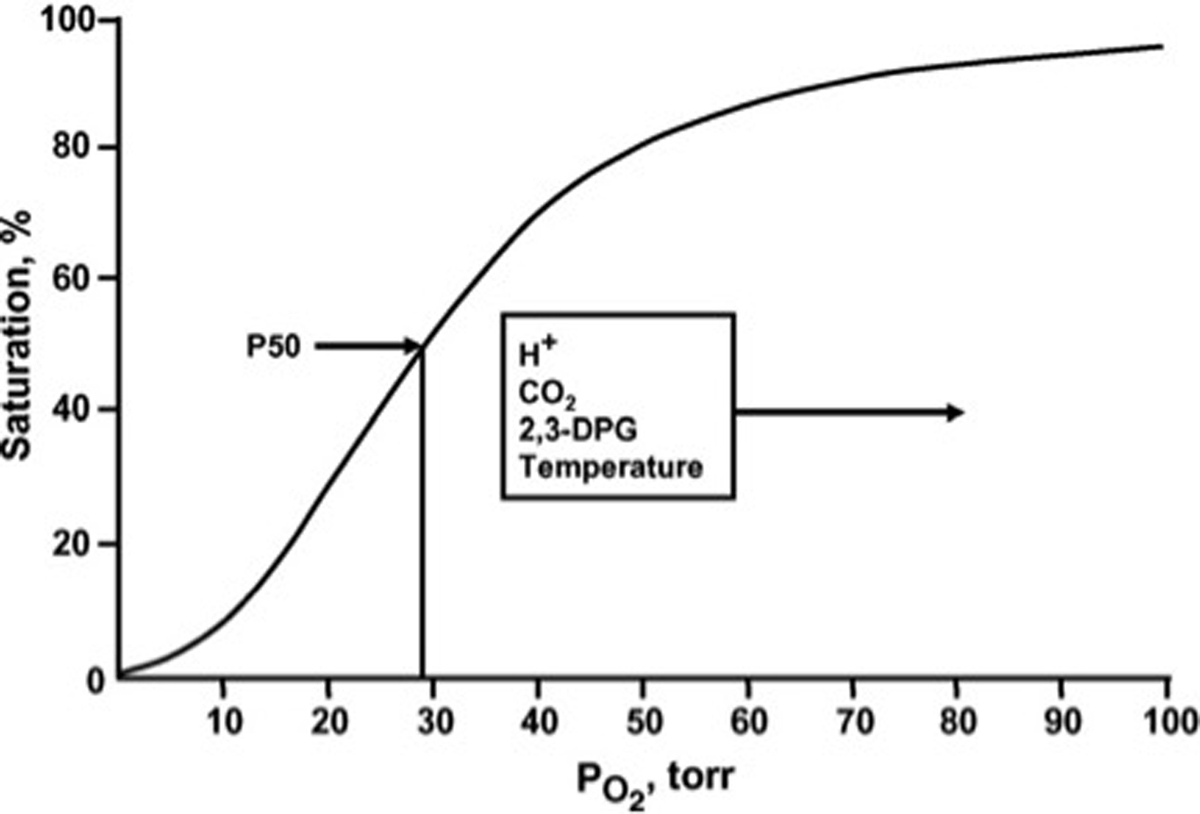
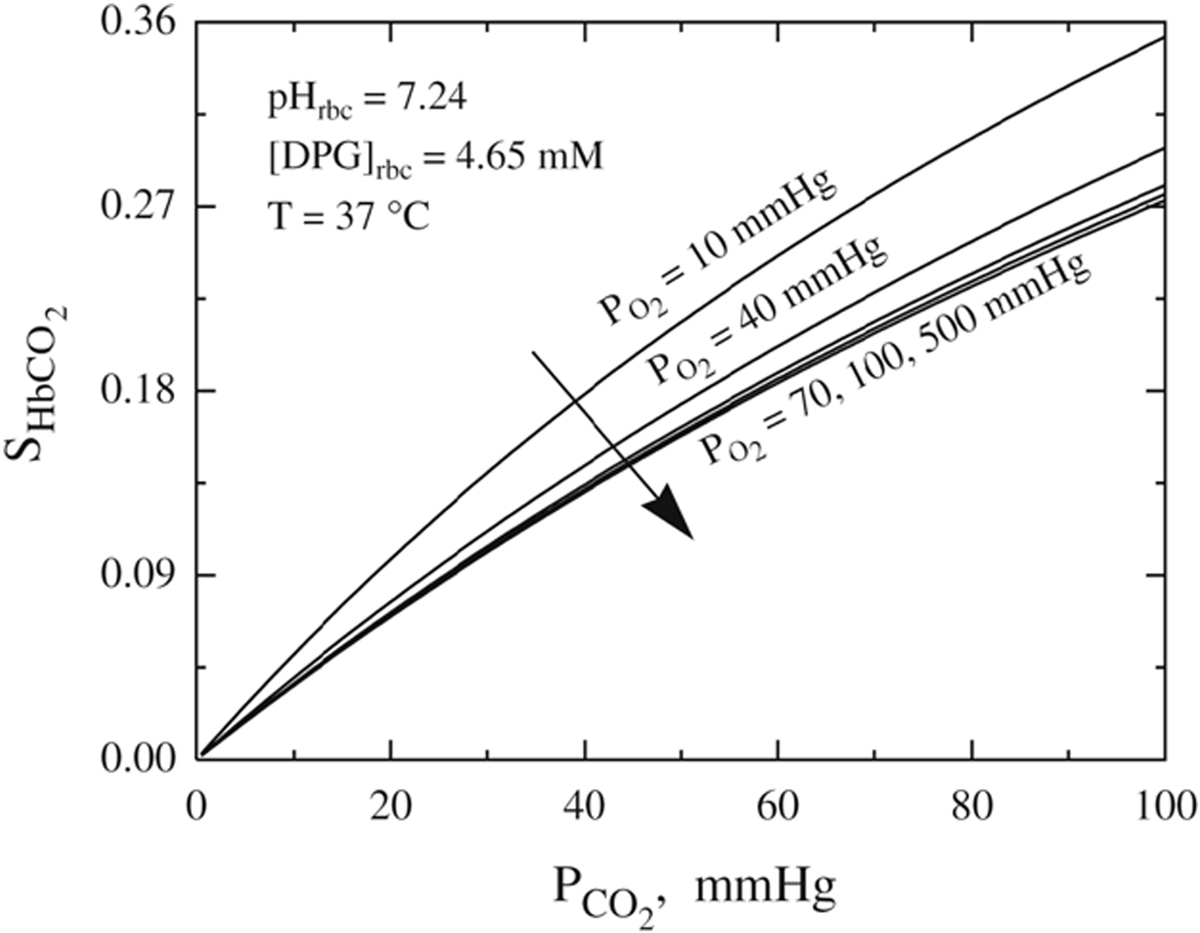
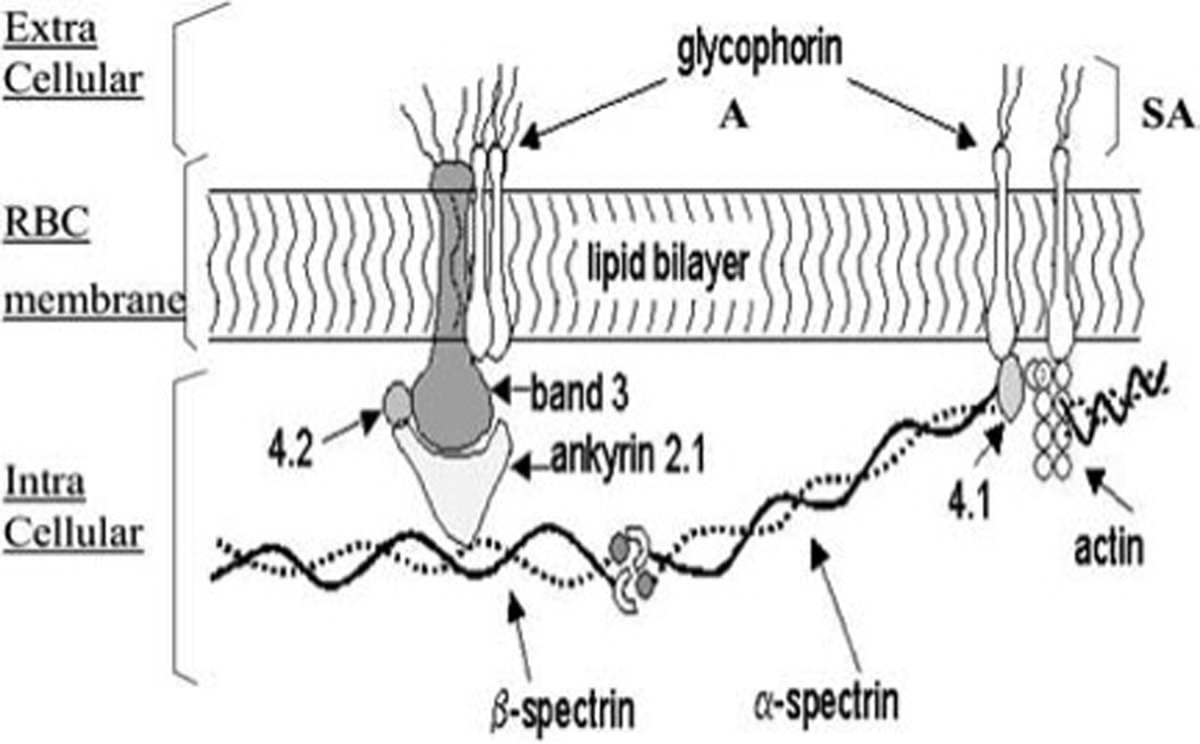
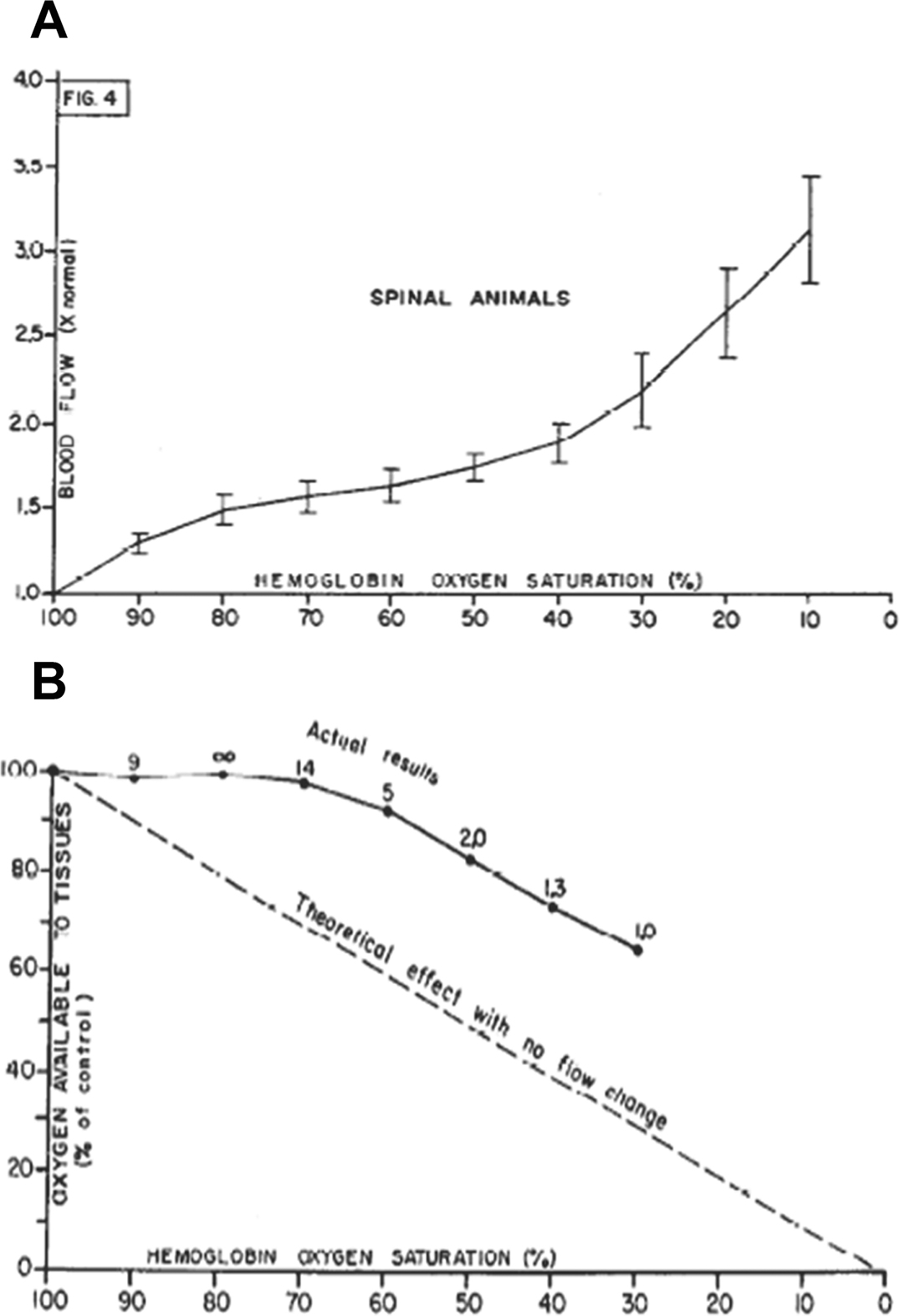
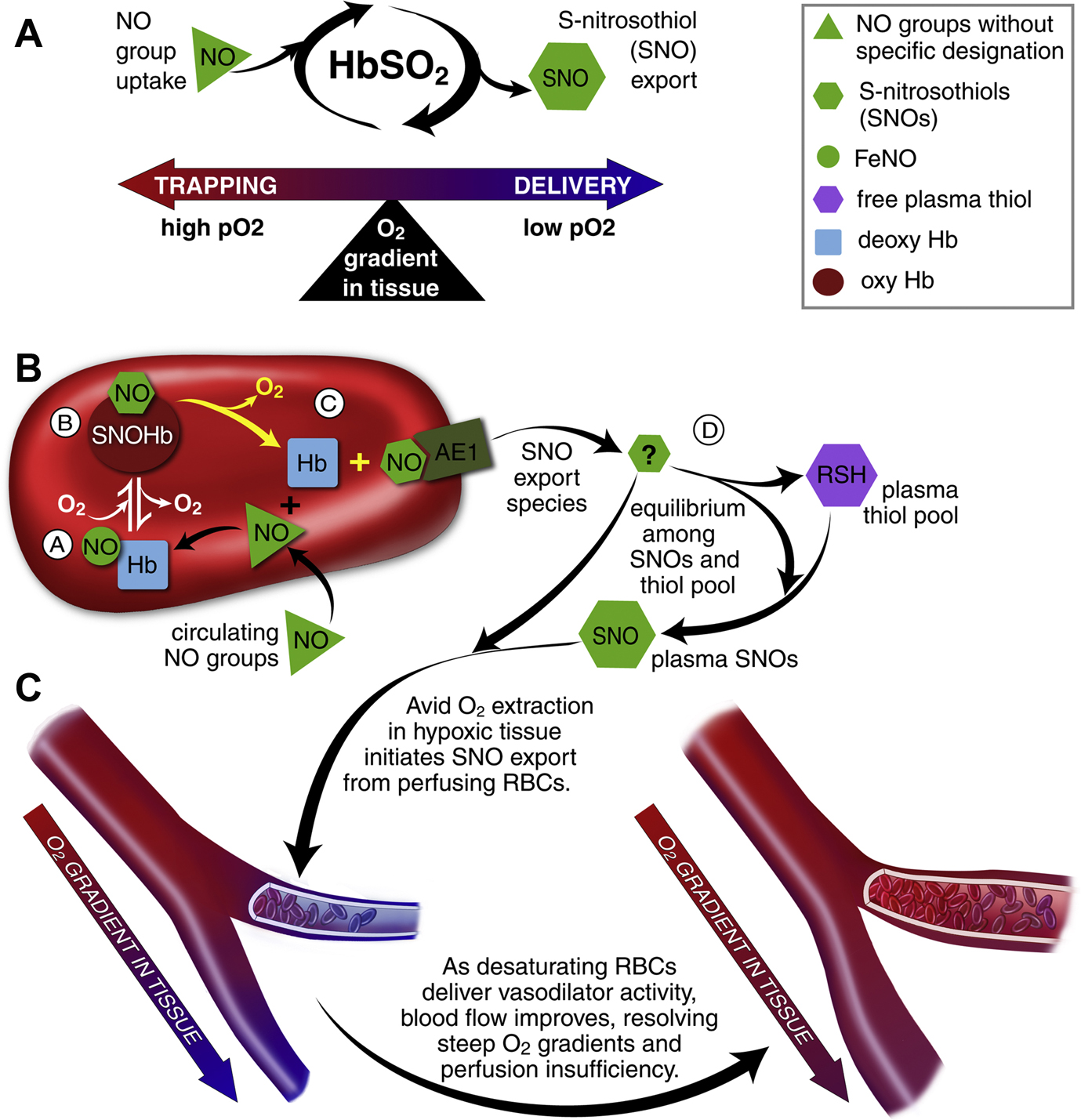
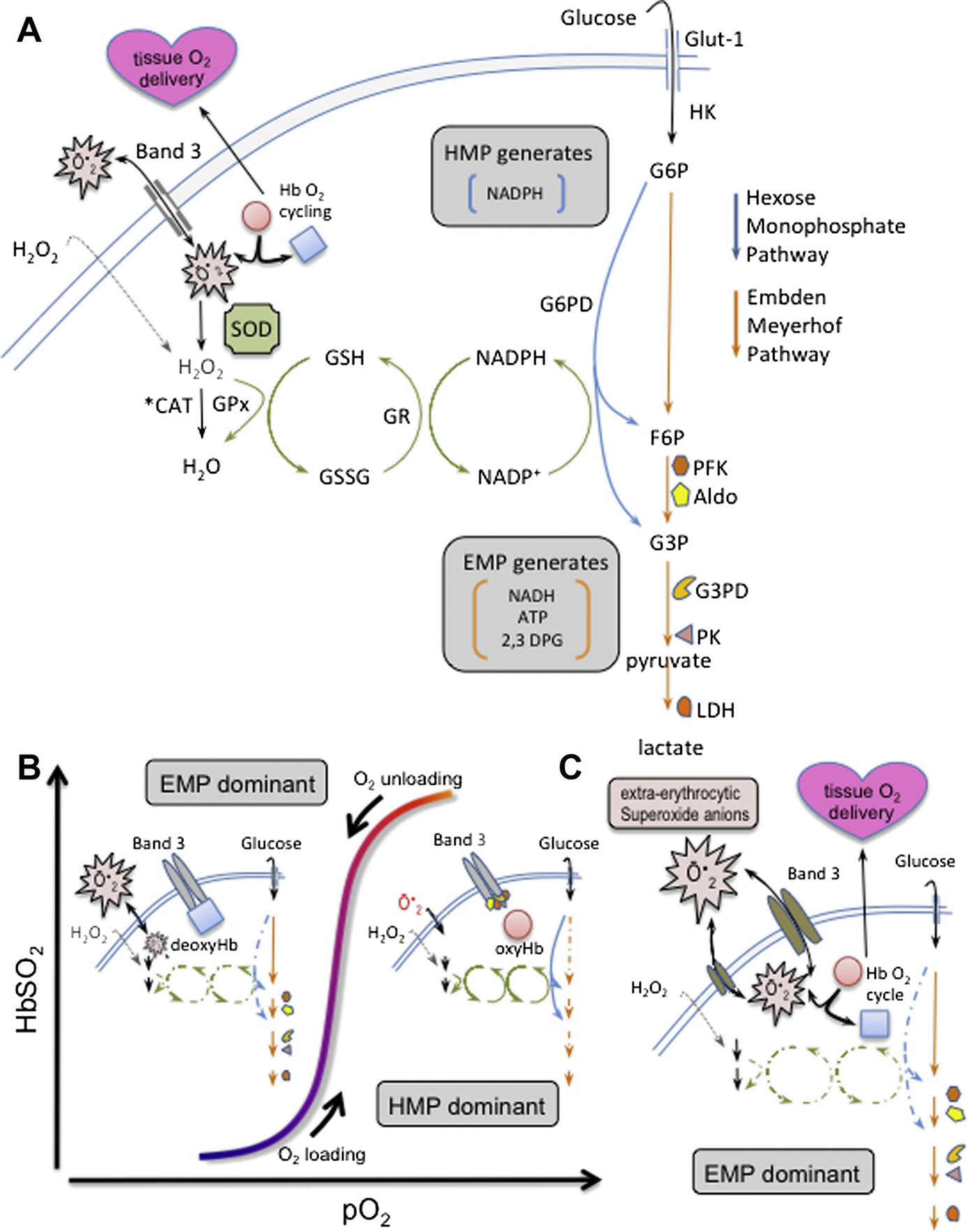
Oxygen (O2) delivery, which is fundamental to supporting patients with critical illness, is a function of blood O2 content and flow. This article reviews red blood cell (RBC) physiology and dysfunction relevant to disordered O2 delivery in the critically ill. Flow is the focus of O2 delivery regulation: O2 content is relatively fixed, whereas flow fluctuates greatly. Thus, blood flow volume and distribution vary to maintain coupling between O2 delivery and demand. This article reviews conventional RBC physiology influencing O2 delivery and introduces a paradigm for O2 delivery homeostasis based on coordinated gas transport and vascular signaling by RBCs.
Keywords: Blood flow; Erythrocyte; O(2) delivery; Red blood cell; Vasoregulation.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflicts of Interest A. Doctor has received research funding and/or consulting fees from Viasys Inc, Entegrion Inc, Terumo BCT, Fresenius Kabi, Galleon Pharmaceuticals, Nitrox LLD, Nitric BioTherapeutics, Galera Inc, and Novartis. A. Doctor holds intellectual property related to biosynthetic artificial RBCs and holds equity in and is CSO of KaloCyte, Inc, which is developing this technology. S. Rogers has no conflicts to declare.
Figures
Similar articles
-
Red cell physiology and signaling relevant to the critical care setting.Curr Opin Pediatr. 2015 Jun;27(3):267-76. doi: 10.1097/MOP.0000000000000225. Curr Opin Pediatr. 2015. PMID: 25888155 Free PMC article. Review.
-
Chemical physiology of blood flow regulation by red blood cells: the role of nitric oxide and S-nitrosohemoglobin.Annu Rev Physiol. 2005;67:99-145. doi: 10.1146/annurev.physiol.67.060603.090918. Annu Rev Physiol. 2005. PMID: 15709954 Review.
-
Nitric oxide in red blood cell adaptation to hypoxia.Acta Biochim Biophys Sin (Shanghai). 2018 Jul 1;50(7):621-634. doi: 10.1093/abbs/gmy055. Acta Biochim Biophys Sin (Shanghai). 2018. PMID: 29860301 Review.
-
Sepsis impairs microvascular autoregulation and delays capillary response within hypoxic capillaries.Crit Care. 2015 Nov 5;19:389. doi: 10.1186/s13054-015-1102-7. Crit Care. 2015. PMID: 26537126 Free PMC article.
-
Red blood cell rheology in sepsis.Intensive Care Med. 2003 Jul;29(7):1052-61. doi: 10.1007/s00134-003-1783-2. Epub 2003 Jun 12. Intensive Care Med. 2003. PMID: 12802488 Review.
Cited by
-
The ultimate tradeoff: how red cell adaptations to malaria alter the host response during critical illness.Am J Physiol Lung Cell Mol Physiol. 2023 Feb 1;324(2):L169-L178. doi: 10.1152/ajplung.00127.2022. Epub 2023 Jan 3. Am J Physiol Lung Cell Mol Physiol. 2023. PMID: 36594846 Free PMC article. Review.
-
Red blood cell transfusions post diagnosis of necrotizing enterocolitis and the deterioration of necrotizing enterocolitis in full-term and near-term infants: a propensity score adjustment retrospective cohort study.BMC Pediatr. 2022 Apr 15;22(1):211. doi: 10.1186/s12887-022-03276-4. BMC Pediatr. 2022. PMID: 35428277 Free PMC article.
-
Erythrocytes Functionality in SARS-CoV-2 Infection: Potential Link with Alzheimer's Disease.Int J Mol Sci. 2023 Mar 17;24(6):5739. doi: 10.3390/ijms24065739. Int J Mol Sci. 2023. PMID: 36982809 Free PMC article. Review.
-
The Effect of the Acid-Base Imbalance on the Shape and Structure of Red Blood Cells.Cells. 2024 Nov 3;13(21):1813. doi: 10.3390/cells13211813. Cells. 2024. PMID: 39513920 Free PMC article.
-
Two-centers machine learning analysis for predicting acid-fast bacilli results in tuberculosis sputum tests.J Clin Tuberc Other Mycobact Dis. 2025 Jan 27;38:100511. doi: 10.1016/j.jctube.2025.100511. eCollection 2025 Feb. J Clin Tuberc Other Mycobact Dis. 2025. PMID: 39927134 Free PMC article.
References
-
- Kaushansky K Williams Hematology. 8e ed: McGraw-Hill; 2010.
-
- Doctor A, Stamler JS. Nitric oxide transport in blood: a third gas in the respiratory cycle. Compr Physiol. 2011;1(1):541–568. - PubMed
-
- Hsia CC. Respiratory function of hemoglobin. N Engl J Med. 1998;338(4):239–247. - PubMed
-
- Edsall JT. Understanding blood and hemoglobin: an example of international relations in science. Perspectives in biology and medicine. 1986;29(3 Pt 2):S107–123. - PubMed
-
- Edsall JT. Hemoglobin and the origins of the concept of allosterism. Federation proceedings. 1980;39(2):226–235. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
