

Microglia depletion and alcohol: Transcriptome and behavioral profiles
- PMID: 32176824
- PMCID: PMC8510547
- DOI: 10.1111/adb.12889
Microglia depletion and alcohol: Transcriptome and behavioral profiles
Abstract
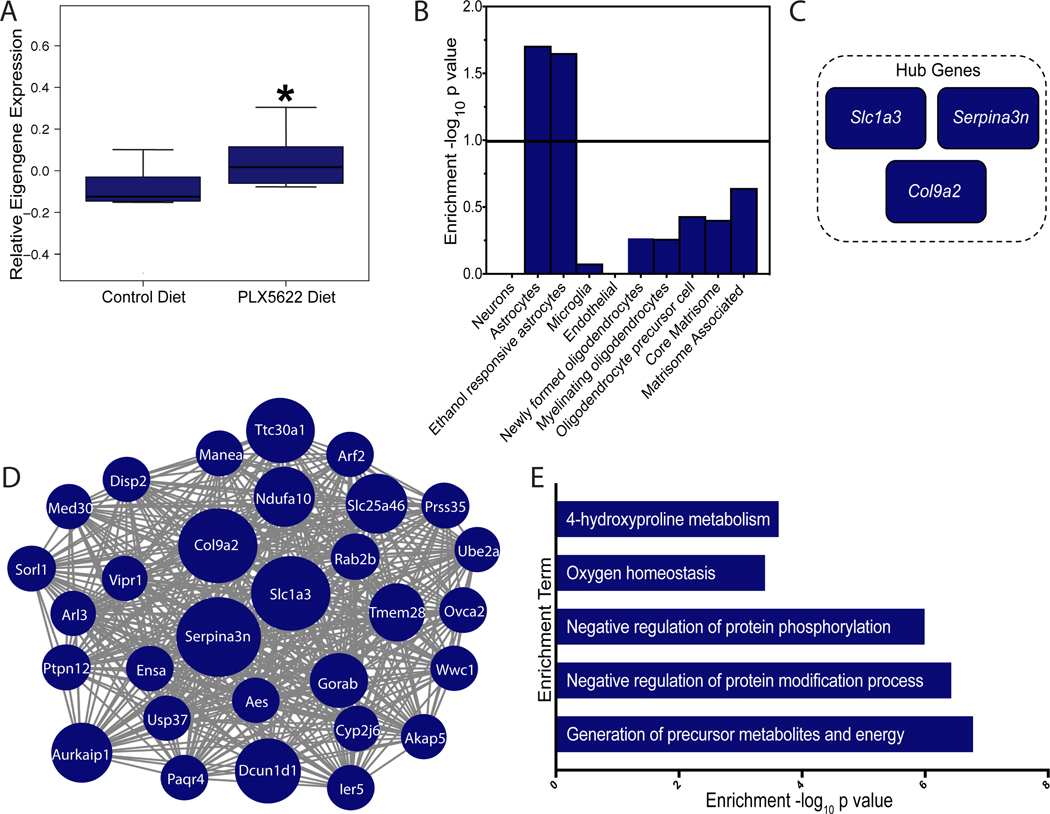
Alcohol abuse induces changes in microglia morphology and immune function, but whether microglia initiate or simply amplify the harmful effects of alcohol exposure is still a matter of debate. Here, we determine microglia function in acute and voluntary drinking behaviors using a colony-stimulating factor 1 receptor inhibitor (PLX5622). We show that microglia depletion does not alter the sedative or hypnotic effects of acute intoxication. Microglia depletion also does not change the escalation or maintenance of chronic voluntary alcohol consumption. Transcriptomic analysis revealed that although many immune genes have been implicated in alcohol abuse, downregulation of microglia genes does not necessitate changes in alcohol intake. Instead, microglia depletion and chronic alcohol result in compensatory upregulation of alcohol-responsive, reactive astrocyte genes, indicating astrocytes may play a role in regulation of these alcohol behaviors. Taken together, our behavioral and transcriptional data indicate that microglia are not the primary effector cell responsible for regulation of acute and voluntary alcohol behaviors. Because microglia depletion did not regulate acute or voluntary alcohol behaviors, we hypothesized that these doses were insufficient to activate microglia and recruit them to an effector phenotype. Therefore, we used a model of repeated immune activation using polyinosinic:polycytidylic acid (poly(I:C)) to activate microglia. Microglia depletion blocked poly(I:C)-induced escalations in alcohol intake, indicating microglia regulate drinking behaviors with sufficient immune activation. By testing the functional role of microglia in alcohol behaviors, we provide insight into when microglia are causal and when they are consequential for the transition from alcohol use to dependence.
Keywords: PLX5622; alcohol; astrocytes; microglia; neuroimmune; transcriptome.
© 2020 Society for the Study of Addiction.
Conflict of interest statement
CONFLICT OF INTEREST
The authors report no biomedical financial interests or potential conflicts of interest.
Figures






References
-
- Li Q, Barres BA. Microglia and macrophages in brain homeostasis and disease. Nat Rev Immunol. 2018;18(4):225–242. - PubMed
-
- Henriques JF, Portugal CC, Canedo T, Relvas JB, Summavielle T, Socodato R. Microglia and alcohol meet at the crossroads: microglia as critical modulators of alcohol neurotoxicity. Toxicol Lett. 2018;283:21–31. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
