

ENO regulates tomato fruit size through the floral meristem development network
- PMID: 32179669
- PMCID: PMC7148573
- DOI: 10.1073/pnas.1913688117
ENO regulates tomato fruit size through the floral meristem development network
Abstract
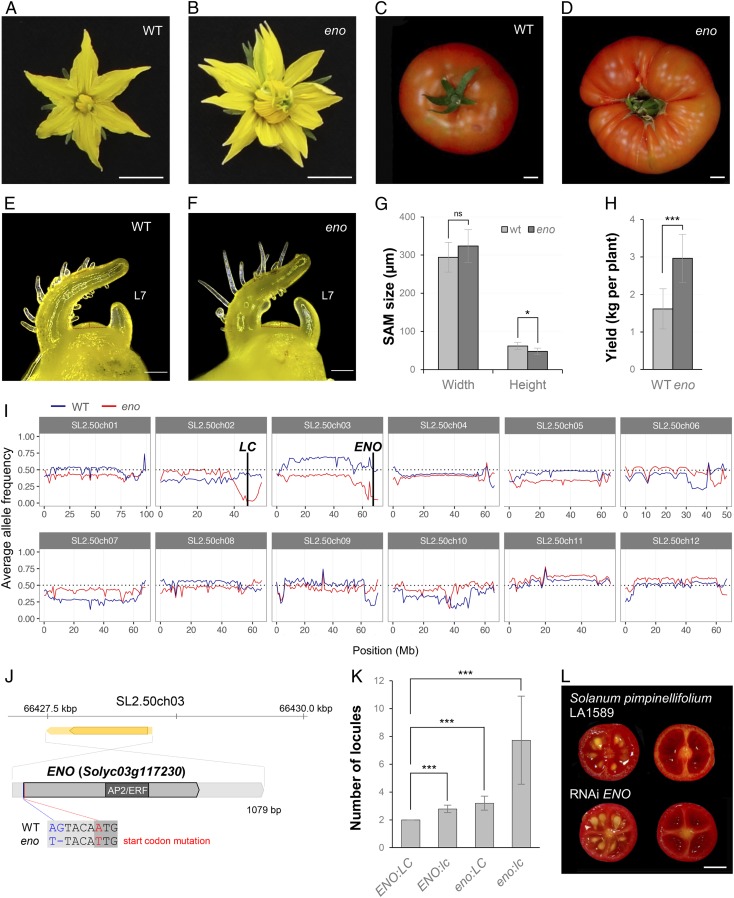
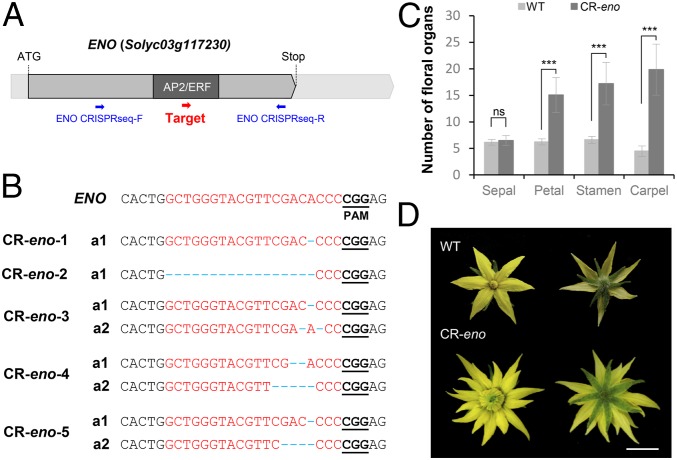
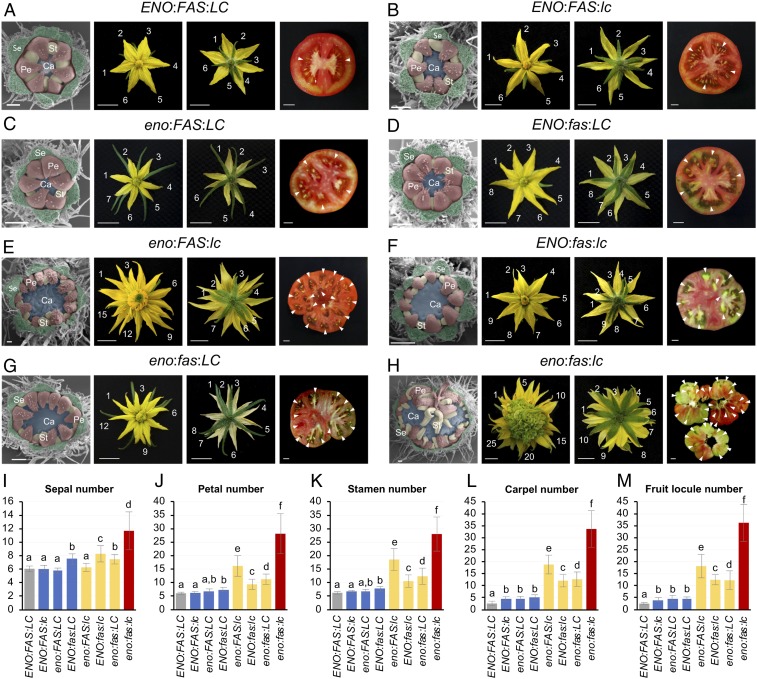
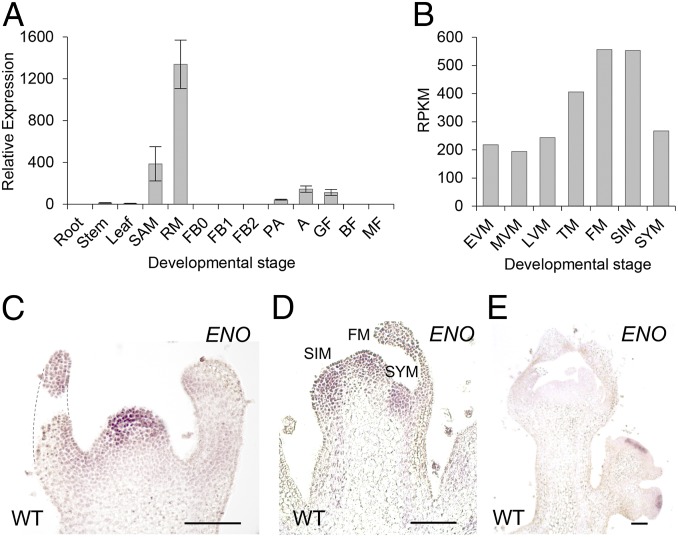
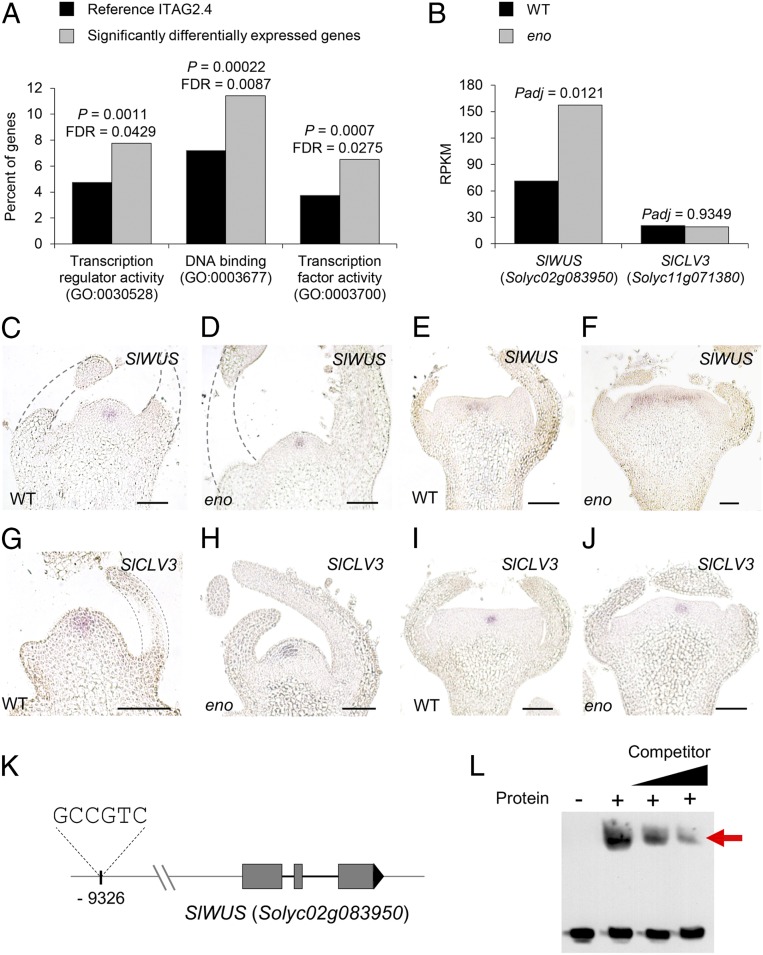
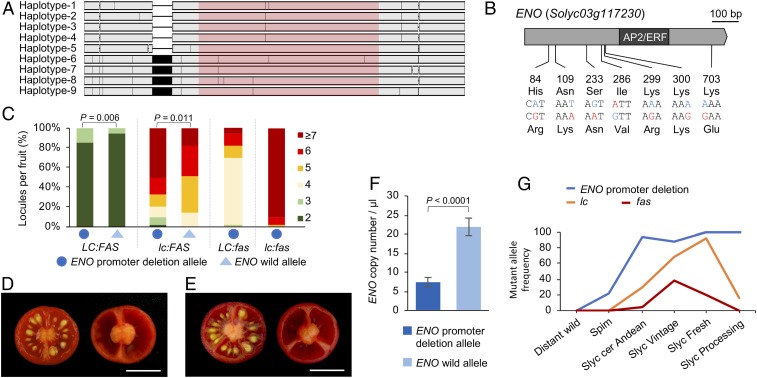
A dramatic evolution of fruit size has accompanied the domestication and improvement of fruit-bearing crop species. In tomato (Solanum lycopersicum), naturally occurring cis-regulatory mutations in the genes of the CLAVATA-WUSCHEL signaling pathway have led to a significant increase in fruit size generating enlarged meristems that lead to flowers with extra organs and bigger fruits. In this work, by combining mapping-by-sequencing and CRISPR/Cas9 genome editing methods, we isolated EXCESSIVE NUMBER OF FLORAL ORGANS (ENO), an AP2/ERF transcription factor which regulates floral meristem activity. Thus, the ENO gene mutation gives rise to plants that yield larger multilocular fruits due to an increased size of the floral meristem. Genetic analyses indicate that eno exhibits synergistic effects with mutations at the LOCULE NUMBER (encoding SlWUS) and FASCIATED (encoding SlCLV3) loci, two central players in the evolution of fruit size in the domestication of cultivated tomatoes. Our findings reveal that an eno mutation causes a substantial expansion of SlWUS expression domains in a flower-specific manner. In vitro binding results show that ENO is able to interact with the GGC-box cis-regulatory element within the SlWUS promoter region, suggesting that ENO directly regulates SlWUS expression domains to maintain floral stem-cell homeostasis. Furthermore, the study of natural allelic variation of the ENO locus proved that a cis-regulatory mutation in the promoter of ENO had been targeted by positive selection during the domestication process, setting up the background for significant increases in fruit locule number and fruit size in modern tomatoes.
Keywords: AP2/ERF transcription factor; CLAVATA-WUSCHEL regulatory network; Solanum lycopersicum; floral meristem; fruit size.
Copyright © 2020 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
Mutation at the tomato excessive number of floral organs (ENO) locus impairs floral meristem development, thus promoting an increased number of floral organs and fruit size.Plant Sci. 2015 Mar;232:41-8. doi: 10.1016/j.plantsci.2014.12.007. Epub 2014 Dec 15. Plant Sci. 2015. PMID: 25617322
-
Developmental characterization of the fasciated locus and mapping of Arabidopsis candidate genes involved in the control of floral meristem size and carpel number in tomato.Genome. 2006 Aug;49(8):991-1006. doi: 10.1139/g06-059. Genome. 2006. PMID: 17036074
-
Meristem activity during flower and ovule development in tomato is controlled by the mini zinc finger gene INHIBITOR OF MERISTEM ACTIVITY.Plant J. 2008 Aug;55(3):415-27. doi: 10.1111/j.1365-313X.2008.03520.x. Epub 2008 Apr 12. Plant J. 2008. PMID: 18410478
-
The genetic basis of fruit morphology in horticultural crops: lessons from tomato and melon.J Exp Bot. 2014 Aug;65(16):4625-37. doi: 10.1093/jxb/eru017. Epub 2014 Feb 11. J Exp Bot. 2014. PMID: 24520021 Review.
-
The CLV-WUS Stem Cell Signaling Pathway: A Roadmap to Crop Yield Optimization.Plants (Basel). 2018 Oct 19;7(4):87. doi: 10.3390/plants7040087. Plants (Basel). 2018. PMID: 30347700 Free PMC article. Review.
Cited by
-
Tomato CRABS CLAW paralogues interact with chromatin remodelling factors to mediate carpel development and floral determinacy.New Phytol. 2022 May;234(3):1059-1074. doi: 10.1111/nph.18034. Epub 2022 Mar 3. New Phytol. 2022. PMID: 35170044 Free PMC article.
-
Recent advances and current challenges in population genomics of structural variation in animals and plants.Front Genet. 2022 Nov 29;13:1060898. doi: 10.3389/fgene.2022.1060898. eCollection 2022. Front Genet. 2022. PMID: 36523759 Free PMC article. Review.
-
CRISPR-Cas9 based molecular breeding in crop plants: a review.Mol Biol Rep. 2024 Jan 28;51(1):227. doi: 10.1007/s11033-023-09086-w. Mol Biol Rep. 2024. PMID: 38281301 Review.
-
CmFUL1 was potentially involved in fruit elongation in melon.Hortic Res. 2025 May 21;12(8):uhaf138. doi: 10.1093/hr/uhaf138. eCollection 2025 Aug. Hortic Res. 2025. PMID: 40698194 Free PMC article.
-
CRISPR/Cas systems: opportunities and challenges for crop breeding.Plant Cell Rep. 2021 Jun;40(6):979-998. doi: 10.1007/s00299-021-02708-2. Epub 2021 May 11. Plant Cell Rep. 2021. PMID: 33977326 Review.
References
-
- Bommert P., Nagasawa N. S., Jackson D., Quantitative variation in maize kernel row number is controlled by the FASCIATED EAR2 locus. Nat. Genet. 45, 334–337 (2013). - PubMed
-
- Xu C., et al. , A cascade of arabinosyltransferases controls shoot meristem size in tomato. Nat. Genet. 47, 784–792 (2015). - PubMed
-
- Je B. I., et al. , Signaling from maize organ primordia via FASCIATED EAR3 regulates stem cell proliferation and yield traits. Nat. Genet. 48, 785–791 (2016). - PubMed
-
- Schoof H., et al. , The stem cell population of Arabidopsis shoot meristems is maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell 100, 635–644 (2000). - PubMed
-
- Somssich M., Je B. I., Simon R., Jackson D., CLAVATA-WUSCHEL signaling in the shoot meristem. Development 143, 3238–3248 (2016). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
