

RUNX1 and RUNX2 transcription factors function in opposing roles to regulate breast cancer stem cells
- PMID: 32180230
- PMCID: PMC7415511
- DOI: 10.1002/jcp.29625
RUNX1 and RUNX2 transcription factors function in opposing roles to regulate breast cancer stem cells
Abstract
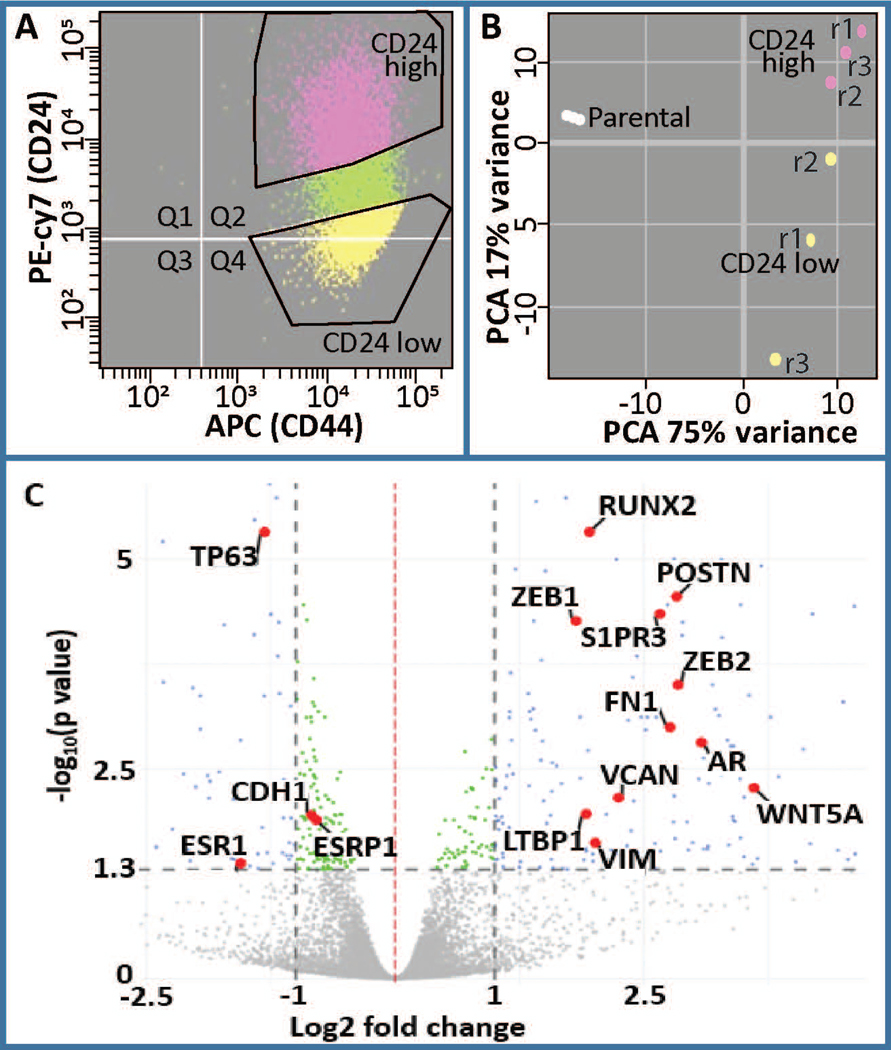
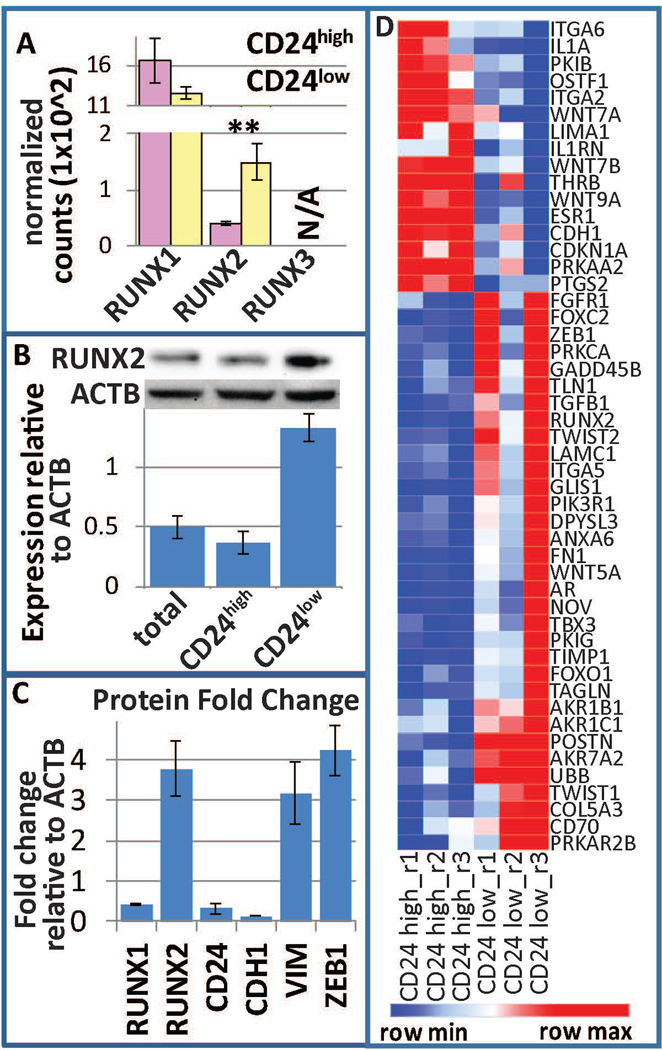
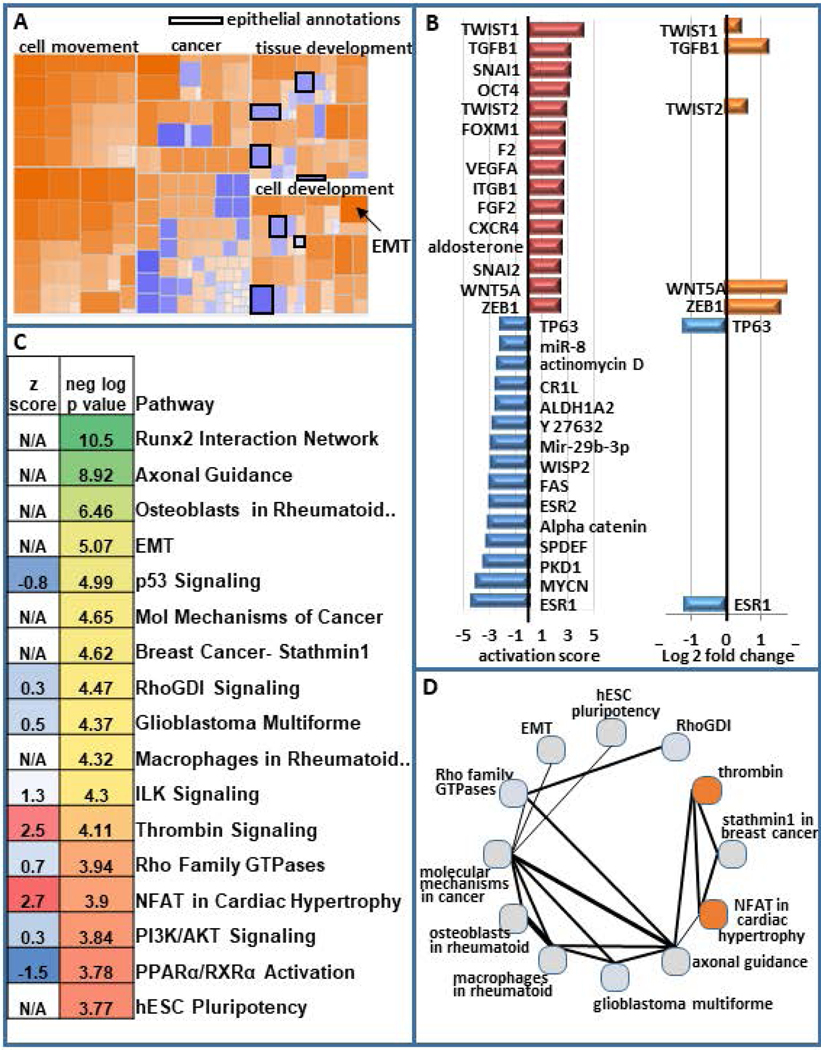
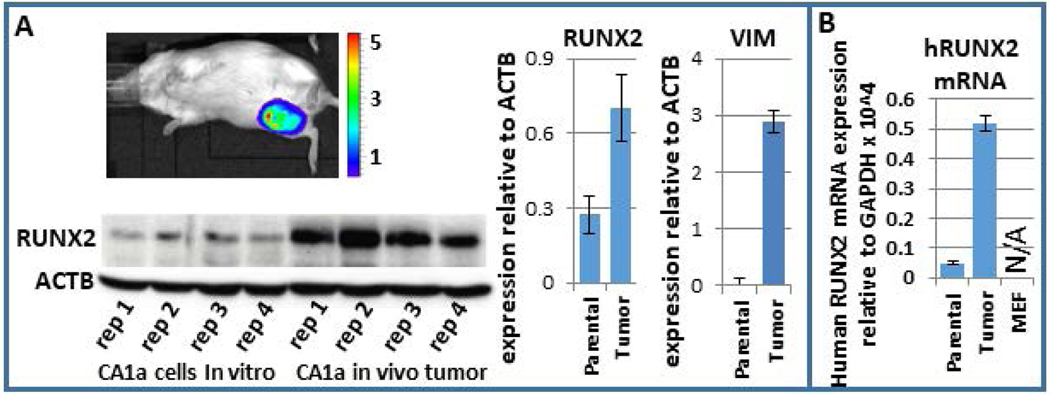
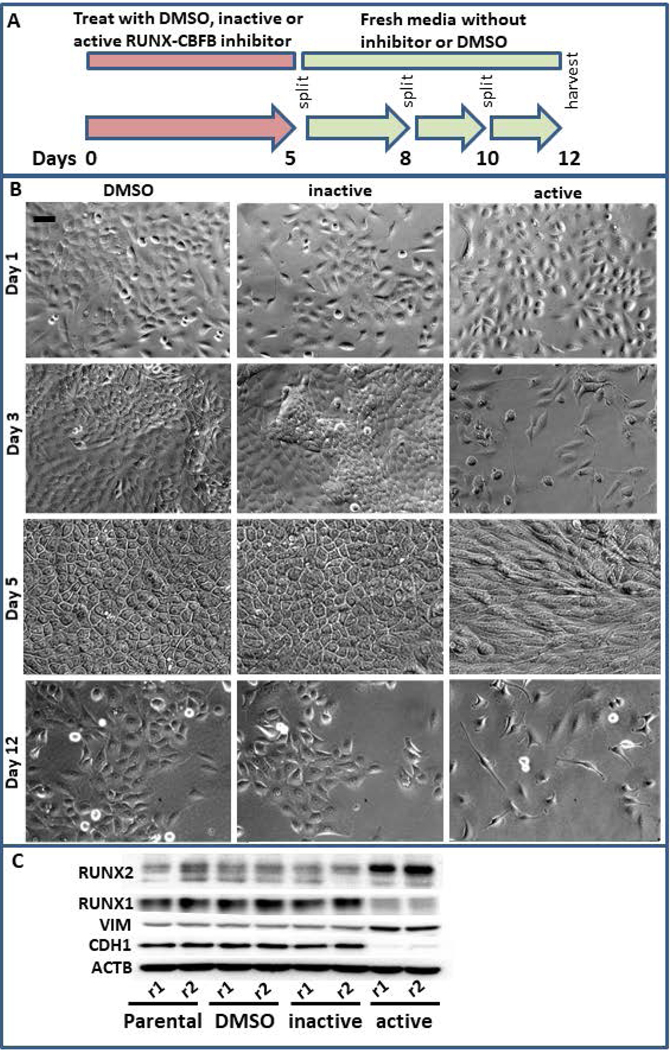
Breast cancer stem cells (BCSCs) are competent to initiate tumor formation and growth and refractory to conventional therapies. Consequently BCSCs are implicated in tumor recurrence. Many signaling cascades associated with BCSCs are critical for epithelial-to-mesenchymal transition (EMT). We developed a model system to mechanistically examine BCSCs in basal-like breast cancer using MCF10AT1 FACS sorted for CD24 (negative/low in BCSCs) and CD44 (positive/high in BCSCs). Ingenuity Pathway Analysis comparing RNA-seq on the CD24-/low versus CD24+/high MCF10AT1 indicates that the top activated upstream regulators include TWIST1, TGFβ1, OCT4, and other factors known to be increased in BCSCs and during EMT. The top inhibited upstream regulators include ESR1, TP63, and FAS. Consistent with our results, many genes previously demonstrated to be regulated by RUNX factors are altered in BCSCs. The RUNX2 interaction network is the top significant pathway altered between CD24-/low and CD24+/high MCF10AT1. RUNX1 is higher in expression at the RNA level than RUNX2. RUNX3 is not expressed. While, human-specific quantitative polymerase chain reaction primers demonstrate that RUNX1 and CDH1 decrease in human MCF10CA1a cells that have grown tumors within the murine mammary fat pad microenvironment, RUNX2 and VIM increase. Treatment with an inhibitor of RUNX binding to CBFβ for 5 days followed by a 7-day recovery period results in EMT suggesting that loss of RUNX1, rather than increase in RUNX2, is a driver of EMT in early stage breast cancer. Increased understanding of RUNX regulation on BCSCs and EMT will provide novel insight into therapeutic strategies to prevent recurrence.
Keywords: RUNX1; RUNX2; breast cancer; breast cancer stem cells; epithelial to mesenchymal transition.
© 2020 Wiley Periodicals, Inc.
Conflict of interest statement
Conflict of Interest
The authors have declared that no conflict of interest exists.
Figures
References
-
- Akech J, Wixted JJ, Bedard K, van der Deen M, Hussain S, Guise TA, . . . Lian JB (2010). Runx2 association with progression of prostate cancer in patients: mechanisms mediating bone osteolysis and osteoblastic metastatic lesions. Oncogene, 29(6), 811–821. doi: 10.1038/onc.2009.389 - DOI - PMC - PubMed
-
- Banerji S, Cibulskis K, Rangel-Escareno C, Brown KK, Carter SL, Frederick AM, . . . Meyerson M (2012). Sequence analysis of mutations and translocations across breast cancer subtypes. Nature, 486(7403), 405–409. doi:http://www.nature.com/nature/journal/v486/n7403/abs/nature11154.html#sup... - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
