

Mechanisms of insulin resistance related to white, beige, and brown adipocytes
- PMID: 32180558
- PMCID: PMC6997501
- DOI: 10.1016/j.molmet.2019.12.014
Mechanisms of insulin resistance related to white, beige, and brown adipocytes
Abstract
Background: The diminished glucose lowering effect of insulin in obesity, called "insulin resistance," is associated with glucose intolerance, type 2 diabetes, and other serious maladies. Many publications on this topic have suggested numerous hypotheses on the molecular and cellular disruptions that contribute to the syndrome. However, significant uncertainty remains on the mechanisms of its initiation and long-term maintenance.
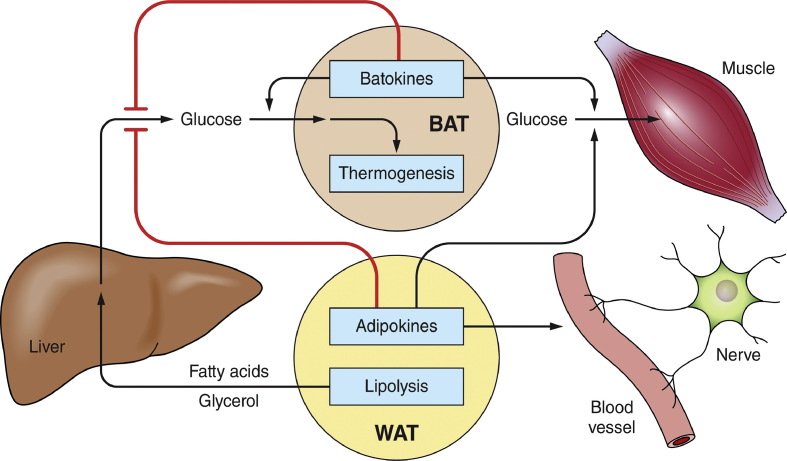
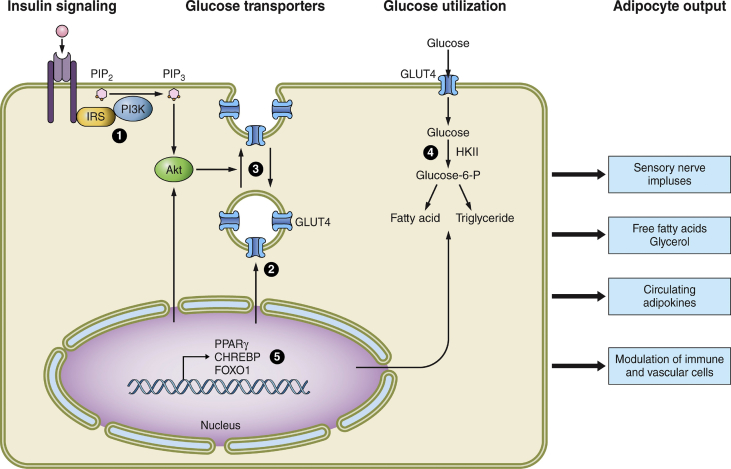
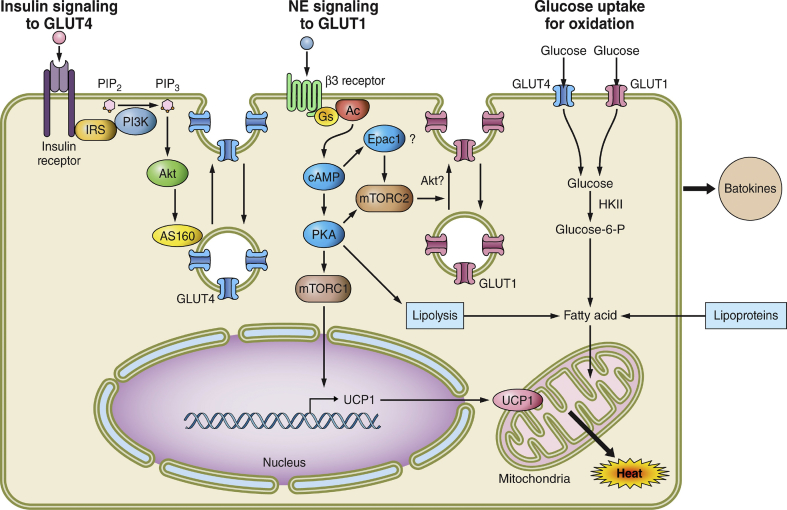
Scope of review: To simplify insulin resistance analysis, this review focuses on the unifying concept that adipose tissue is a central regulator of systemic glucose homeostasis by controlling liver and skeletal muscle metabolism. Key aspects of adipose function related to insulin resistance reviewed are: 1) the modes by which specific adipose tissues control hepatic glucose output and systemic glucose disposal, 2) recently acquired understanding of the underlying mechanisms of these modes of regulation, and 3) the steps in these pathways adversely affected by obesity that cause insulin resistance.
Major conclusions: Adipocyte heterogeneity is required to mediate the multiple pathways that control systemic glucose tolerance. White adipocytes specialize in sequestering triglycerides away from the liver, muscle, and other tissues to limit toxicity. In contrast, brown/beige adipocytes are very active in directly taking up glucose in response to β adrenergic signaling and insulin and enhancing energy expenditure. Nonetheless, white, beige, and brown adipocytes all share the common feature of secreting factors and possibly exosomes that act on distant tissues to control glucose homeostasis. Obesity exerts deleterious effects on each of these adipocyte functions to cause insulin resistance.
Keywords: Adipokines; Adipose tissues; Adrenergic receptors; Bioactive lipids; Glucose tolerance; Lipogenesis; Signaling; Thermogenesis; Uncoupling protein.
Copyright © 2020 The Author. Published by Elsevier GmbH.. All rights reserved.
Figures
References
-
- Marks V., Richmond C. 1st ed. Royal Society of Medicine; London: 2007. Insulin murders.
-
- Sweeney J.S. Dietary factors that influence the dextrose tolerance test. A Preliminary Study. Archives of Internal Medicine. 1927;40(6):818–830.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
